
Attenuation of Microcystins Using Electron Beams and Gamma Radiation: A Study with Environment-Bound Conditions
Abstract
Microcystins (MCs) are toxins profusely synthesized by cyanobacteria, causing livestock poisonings and endangering human health. We design and execute an experiment to investigate the attenuation (degradation) of microcystins by exposing them to gamma radiation and electron beams at doses of 0 (control), 3, 5, 10 and 15 kGy. The experimental conditions simulate microcystin contamination of aquatic environments; we thus consider (1) microcystins inside whole cells and extracellular dissolved in water, simulated by non-sonicated and sonicated cells, respectively, and (2) two acute microcystin concentrations within water. Toxicity tests of Microcystis aeruginosa detected immobilization (i.e., paralysis) of Ceriodaphniasilvestrii exposed to aqueous crude extracts of irradiated and non-irradiated M. aeruginosa (NPLJ-4 strain) at concentrations of 45 and 90 mg.L-1 (mg dry weight of freeze-dried material), and the results were analyzed using the Trimmed Spearman-Karber statistical program to obtain 48-h EC50, the average effective concentration causing immobility in 50% of organisms after 48 hours.
We conclude that electron beams are effective physical agents for toxin attenuation (degradation) and reach 100% effectiveness at 5 kGy and above; their efficiency is two orders of magnitude greater than that of gamma radiation.
This new body of information contributes to (1) remediating environmental water sources; (2) designing water/wastewater treatment facilities; (3) combatting chronic microcystin environmental contamination; and (4) inspiring further studies to promote the use of biomonitors (e.g., Cladocerans) to detect and evaluate microalgae contamination.
Author Contributions
Academic Editor: Wei Wu, Nanjing Medical University.
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2021 Joao Dias Toledo Arruda-Neto, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Competing interests
The authors have declared that no competing interests exist.
Citation:
Introduction
Toxic cyanobacteria blooms in surface waters are a major worldwide environmental concern. They produce bioactive molecules such as microcystins (MC), some of which produce cyanotoxins 1, 2. Of all microcystin variants, microcystin-LR (MC-LR) predominates. Microcystins cause poisonings in livestock and wildlife worldwide and threaten human health by contaminating drinking water and recreational waters 3, 4. Study author M. C. Bittencourt-Oliveira has researched micro cystin-related food poisoning by studying the photosynthetic effects and bioaccumulation of microcystin congeners in lettuce irrigated with contaminated water 5.
Microcystins have been detected dissolved in the water of reservoirs, notably the Guarapiranga dam in São Paulo City, Brazil, which supplies water to 25% of the city’s population. Our laboratory previously studied heavy metal contamination in this reservoir and how it affects the surrounding population 6, 7.
A well-documented microcystin exposure accident occurred in 1997 at a haemodialysis center in Brazil and was caused by microcystin-producing cyanobacteria, detected in reservoir water supplying one of the dialysis centers. Fifty out of one hundred patients died, and to date this is the most serious microcystin poisoning in humans worldwide 8.
A comprehensive review by Franca Buratti and associates discusses numerous aspects of cyanotoxins, such as their removal in water treatment plants, occurrence, toxicity, and risk evaluation 9. Conventional treatments (coagulation, flocculation, sedimentation and filtration) are somewhat effective at removing cyanotoxins from water, but some conventional treatments result in cellular lysis and thus increase the amount of cyanotoxins dissolved in water 10, 11, 12, 13.
Ionizing radiation, such as gamma radiation and electron beams, effectively reduces pathogenic agents 14, 15, 16 and degrades persistent, non-biodegradable pollutants. It is also a promising technique for the attenuation of microcystins, and this promise motivates the present study. Because ionizing electrons and gamma radiation are promising tools, we need detailed studies on biophysical aspects of radiation-induced microcystin attenuation and should design experimental conditions similar to realistic environment contamination scenarios.
Our study thus investigates microcystin attenuation of toxicity embracing the following situations:
Aquatic Environment Scenarios
(1) Our experiments consider intracellular microcystins and extracellular microcystins dissolved in water using non-sonicated and sonicated cells, respectively, and (2) two acute microcystin concentrations in water.
Radiation-Induced Microcystin Attenuation and Monitoring (1) We use an ample interval of doses of 0, 3, 5, 10 and 15 kGy, and (2) bioassays with Cladocerans to evaluate the microcystin toxicity attenuation.
Quantification of Toxin Attenuation
We express the experimental results as an ad hoc attenuation coefficient, as a function of doses imparted to microcystin water solutions at different concentrations and doses.
From this body of new and fuller information on microcystins exposed to electrons and gamma radiation, we contemplate the following environmental aspects: (1) remediation of water sources contaminated by microcystins using ionizing radiation (gammas and electrons), illustrated by a working example; (2) a better conceptual and experimental understanding of the behavior of microcystins as contaminants, and (3) contribution to ameliorate strategies to combat chronic contamination scenarios of the environment.
Materials and Methods
Interaction of Ionizing Radiation with Biological Matter in Water
Radiolysis of Water
The bioassays with toxins are performed in aqueous medium. Exposing toxins in aqueous samples to gamma radiation and electron beams induces the radiolysis of water, where reaction products are mostly free radicals, ions, oxygen peroxide and molecular hydrogen. This gamma-induced-radiolysis is as follows:
H2O + γ → e−aq, ·H,HO•, H+, OH-, H2, H2O2,
where an important example is the hydroxyl radical(HO•), a molecule with one unpaired electron on the oxygen atom. The unpaired valence electron in a free radical, the hydroxyl (HO•), endows it with high reactivity, instability and short life 17, 18.
Exposure to high doses of ionizing radiation results in oxidative stress(OS) and increases reactive oxygen species (ROS), where the most reactive ROS, as peroxyl, leads to oxidation of proteins and DNA, among other damaging processes.
Subtleties of Irradiation: Secondary Electrons
Damaging processes to DNA and large molecules, as e.g. proteins and cyanotoxins, are initiated by direct impact of radiation on water molecules, leading to free radical formation by breaking down chemical bonds. Additionally, incidental radiation gives rise to low-energy secondary electrons (smaller than ≈ 20 eV) at significant fluxes. Approximately 5 × 104 secondary electrons are produced per 1 MeV of transferred energy 19, 20, playing an important role in inducing DNA and protein damage.
Electrons, in turn, are charged particles transferring most of their energies to the aqueous medium molecules, that is, water itself plus larger molecules therein dissolved, thus producing copious fluxes of secondary electrons. In fact, the stopping power of gammas in water is much weaker than that of electrons, explaining why their production of secondary electrons is considerably smaller.
Cyanobacterial Cultures: Preparation for Exposure to Gamma Radiation and Electron Beams
We used the cyanobacteria strain of toxin-producing Microcystis aeruginosa Kutzing 1846, NPLJ-4, and a non-toxic strain of Microcystis sp. supplied by Dr. Armando Vieira at the Federal University of São Carlos, Brazil. Both strains were cultivated in 6 L Erlenmeyer flasks containing 4 L ASM-1 medium 21, with the pH adjusted between 7.0 and 7.5. The strains were maintained under a light:dark photoperiod of 12:12 h and at a constant temperature of 25.0 ± 0.2 °C. The biological material was harvested at the late exponential phase of growth, centrifuged (Quimis-Q 222T) and lyophilized at −80 °C (Lyophilizer LB 5000 TT-TERRONI-FAUVEL) until fully dehydrated.
The crude cyanobacterial extract samples were prepared with 100 mg of freeze-dried cells and 1 L of distilled water (stock solution). Two separate procedures, referred to in this study as “sonicated” and “non-sonicated”, were performed in order to evaluate the degradation of microcystin through ionizing radiation. First the solution was homogenized and centrifuged, and then the toxins were sonicated in aqueous medium, and finally the microcystin in the supernatant was exposed to radiation. After homogenization, the crude extract of the non-sonicated samples (toxin inside the cell) was first directly exposed to radiation and then sonicated and centrifuged.
Both sonicated and non-sonicated samples were irradiated with two types of ionizing radiations: gamma and electron beams at doses of 0 (control), 3, 5, 10 and 15 kGy. Gamma radiation was carried out with a 60Co gamma source facility (Gammabeam, model 650 from MSD Nordion, Ottawa, Canada) at a rate of 1.98 kGy h-1. Irradiation with electron beams (LINAC, model Dynamitron DC 1500/25/4, job 188) was performed with an energy of 1.174 MeV and at the following doses: 3 kGy for 03 min. at 6.70 kGy s-1 dose rate; 5 kGy for 03 min. at 11.16 kGy s-1 dose rate; 10 kGy for 03 min. at 22.32 kGy s-1 dose rate; and 15 kGy for 06 min. at 16.74 kGy s-1 dose rate.
Bioassays
Toxicity tests of M. aeruginosa subjected to gamma radiation were performed with Ceriodaphnia silvestrii (Cladocera, Crustacea). For irradiation with electron beams, bioassays included C. silvestrii and two additional species: Argyrodiaptomus furcatus and Notodiaptomus iheringi (Copepoda, Calanoida).
Stock cultures of C. silvestrii Daday 1902 have been maintained for several years at the Ecotoxicology Laboratory of the Federal University of São Carlos, Brazil, reproducing parthenogenetically. The individuals used in this experimental setup originate from a single female to eliminate variation among clones. Cultures were maintained in 2 L glass beakers, and the medium used (modified from 22) was reconstituted water (pH 7.0–7.6; conductivity 160 mS cm-1 and hardness 40 to 48 mg CaCO3 L-1) and renewed three times weekly. Cladocerans were fed daily with Pseudokirchneriellasubcapitata at a concentration of 3 × 105 cell mL-1 and a yeast suspension (1 mL.L-1) prepared with active commercial dried yeast (Fleischmann’s®) and distilled water (0.25 g in 50 mL-1). Fifty individuals per beaker were kept under a light: dark photoperiod of 12:12 at 25 ± 1 °C in an incubator. All experiments were initiated with third brood neonates (≤ 24 h old) derived from healthy parent stock.
Two week before beginning the experiments, we evaluated the health and sensitivity of the cultures by acute toxicity tests with the reference compound sodium chloride, NaCl 22.
Acute toxicity tests were carried out with C. silvestrii, A. furcatus and N. iheringi to analyze their survival during exposure to aqueous crude extracts of irradiated and non-irradiated Microcystis aeruginosa (NPLJ-4 strain), detecting immobilization (i.e., paralysis) of the organisms exposed. The concentrations are given as milligrams of freeze-dried material per liter of water. The results of the acute toxicity tests with C. silvestrii, for instance, were analyzed using the Trimmed Spearman-Karber statistical program 23, obtaining 48-h EC50, that is, the average effective concentration causing immobility in 50% of the organisms exposed to the toxic agent over a period of 48 hours.
Results
Non-Toxic Strain Irradiated with Ionizing Radiation:
Acute toxicity tests were carried out with sonicated and non-sonicated non-toxic Microcystis sp. irradiated with gamma radiation and electron beams at doses 3, 5, 10 and 15 kGy. None of the samples showed toxicity to Ceriodaphnia silvestrii.
Toxicity Test with Toxic Strain of Microcystis Aeruginosa (NPLJ-4), and Exposure to Ionizing Radiation
All tests performed with the control dose 0 kGy of the toxic culture represented by strain NPLJ-4 showed toxic effects on C. silvestrii, with a mean value of 48-h EC50 equal to 31.4 mg.L-1 dry weight of freeze-dried material and a 95% confidence interval of 26.0–38.0 mg.L-1 dry weight of freeze-dried material (Table 1).
Table 1. Effective Concentration of toxic strain NPLJ 4, exposed to ionizing radiation doses, causing immobility in 50% of C. silvestrii| Dose( kGy ) | 48h-EC 50 * Sonicated Samples ( S. D.) ** | 95%(CI) ( mg.L -1 dry weight of freeze-dried material) | 48h-EC 50 Non-Sonicated Samples (S. D.)* * | 95%(CI) ( mg.L -1 dry weight of freeze-dried material) | p value |
| 0 | 31.4 (6.42) | 26.1–38.0 | - | - | - |
| 3 | 49.8 (8.22) | 43.6–56.9 | 46.3 (7.53) | 40.3–53.3 | 0.56 |
| 5 | 47.9 (7.19) | 41.6–55.1 | 42.6 (3.07) | 44.3–57.6 | 0.25 |
| 10 | 62.9 (2.39) | 54.1–65.2 | 50.5 (7.42) | 36.3–50.1 | 0.02 |
| 15 | 65.4 (0.69) | 59.1–71.8 | 64.3 (1.63) | 59.8–70.8 | 0.88 |
The crude extracts irradiated with doses of 3, 5, 10 and 15 kGy gamma radiation systematically exhibited a higher 48-h EC50 than the non-irradiated crude extracts. Both sonicated and non-sonicated samples reflected a loss of toxicity that may be attributed to efficient degradation of cyanotoxins. At the doses 10 kGy and 15 kGy, sonicated and non-sonicated crude extracts exhibited 48-h EC50 with values higher than the control. Statistical analysis of 48-h EC50 obtained in this study are shown in Table 1 and reveal that, with the exception of irradiation with 10 kGy, there was no difference between non-sonicated and sonicated crude extracts (p = 0.05, Student’s t-test) 24.
For bioassays carried out with sonicated crude extracts and irradiated with electron beams at doses of 3, 5, 10 and 15 kGy, the dose of 3 kGy obtained only 48-h EC50 (70.6 mg.L-1 dry weight of freeze-dried material), and we observed immobility of neonate of C. silvestrii. For non-sonicated crude extract irradiated with electron beams with doses of 3 and 5 kGy, we observed immobility of Ceriodaphniasilvestrii, presenting a 48-h EC50 of 73.9 and 62.2 mg.L-1 dry weight of freeze-dried material, respectively. Results for immobility tests of acute toxicity to Ceriodaphniasilvestrii (Crustacea, Cladocera) following irradiation with gammas and electrons are shown in Table 2.
We believe that attenuation of the toxin occurred for crude extract sonicated and irradiated with 5 kGy due to immobilities occurring in non-significant numbers (i.e., an amount insufficient to obtain 48-h EC50). For non-sonicated crude extract at the same dose, the percentage of immobile neonates was elevated. The percentage of immobile individuals (data not shown in Table 2) in sonicated crude extract exposed to electron beams (toxins in aqueous medium) was 1.5%, and non-sonicated crude extract (toxin inside the cell) was 21.5% of the individuals. This behavior could be attributed to the presence of the membrane during irradiation. In fact, the membrane is an additional mass interposed between the electron beams and the toxin within the cells, and thus the dose effectively imparted to the toxin could be lower. This is not the case in irradiation with gammas due to the high penetrability of this radiation.
Other results from bioassays were obtained from toxins irradiated with electron beams. For instance, Borrely and collaborators 25, using Ceriodaphniadubia, Daphnia similis and Vibrio fischeri as test organisms, observed high effectiveness of electron beams in the degradation of the surfactant dodecyl pbenzene sulphonate acid (LAS). They showed that exposing waste from two wastewater treatment plants to electron beams proved to be highly effective for mitigating toxicity 25. Also, Romanelli and collaborators demonstrated reduced toxicity of the surfactant sodium dodecyl sulfate (SDS) when it was diluted with water and subjected to irradiation with electron beams at doses of 3, 6, 9 and 12 kGy 26. The evaluation was performed via acute toxicity tests on two test organisms, the bacterium Vibrio fischeri and the cladoceran Daphnia similis, finding the dose of 6 kGy to be sufficient in mitigating this highly toxic substance.
Gamma Radiation and Electron Beams
In the treatment of sonicated and non-sonicated crude extracts, we used four replicates, and the concentrations in the ecotoxicity tests with C. silvestrii were 5.7, 11.3, 22.5, 45.0 and 90.0 mg.L-1 dry weight of freeze-dried material. For crude extracts irradiated with both gammas and electron beams, the immobility of neonate C. silvestrii was better observed at the higher concentrations of 45 and 90 mg.L-1 dry weight of freeze-dried material. For a concerted visualization of the results, Table 2 shows total values of C. silvestrii neonate immobilities after acute toxicity tests (48 h). For sonicated crude extracts exposed to 3 kGy of gamma radiation, the number of immobilities was almost three times greater than the value for the crude extracts irradiated with electron beams, at a concentration of 45 mg.L-1 dry weight of freeze-dried material. With this same concentration and at doses of 5, 10 and 15 kGy, we observed immobility only for the crude extracts irradiated with gamma, and no immobility for those crude extracts irradiated with electron beams. At a concentration of 90 mg.L-1 dry weight of freeze-dried material and in crude extracts sonicated and irradiated with gamma radiation, the percentage of immobility of organisms occurred after the acute test (48 h). Immobility of organisms was 100% at doses of 3 and 5 kGy, while for 10 and 15 kGy the value was 98.8% and 90%, respectively. The non-sonicated crude extracts showed a number of neonate immobilities greater than sonicated crude extracts after the acute test (48 h). Immobility of organisms occurred in crude extracts exposed to gamma radiation at doses of 3, 5 and 10 kGy and toxin concentration of 45 mg.L-1. One single immobile individual was observed after 48 hours for acute toxicity tests with C. silvestrii in four replicates, for crude extract irradiated with 15 kGy.
Introducing the Toxin Attenuation Parameters: Aγ and Ae
A better and more concise appraisal of the body of data produced in this work could be achieved through the Toxin Attenuation Parameter concept. By naming Nm and Nimm, the number of mobile and immobile Cladocerans (Ceriodaphniasilvestrii) after exposure to aqueous crude extracts of Microcystis aeruginosa, respectively, the Toxin Attenuation Parameter is given by
Ai(%) = (Nimm/NT)x100, where NT = Nm + Nimm, and (1) i = γ or e, standing for gamma radiation and electron beams, respectively.
For instance, Ai(%) = 0% if Nimm= 0, and Ai(%) = 100% if Nimm = NT.
Figure shows the results for Aγ and Ae calculated from the raw data set, Table 2, summarized in Table 3.
Table 2. Immobility results for C. silvestrii exposed to toxic strain NPLJ 4 exposed to ionizing radiation doses| Crude Extract “ Sonicated ” | Crude Extract “Non-Sonicated” | ||||||||
| Doses ( kGy ) | Concentrations* | Gamma Radiation | % Immobilities | Electron Beams | % Immobilities | Gamma Radiation | % Immobilities | Electron Beams | % Immobilities |
| 0 | 45 | 86/120 | 71.7 | - | - | - | - | - | - |
| 90 | 120/120 | 100 | - | - | - | - | - | - | |
| 3 | 45 | 24/80 | 30 | 09/80 | 11.3 | 38/80 | 47.5 | 0/80 | 0 |
| 90 | 80/80 | 100 | 48/80 | 60 | 80/80 | 100 | 58/80 | 72.5 | |
| 5 | 45 | 34/80 | 42.5 | 00/80 | 0 | 44/80 | 55 | 8/80 | 10 |
| 90 | 80/80 | 100 | 06/80 | 7.5 | 80/80 | 100 | 74/80 | 925 | |
| 10 | 45 | 02/80 | 2.5 | 00/80 | 0 | 28/80 | 35 | 0/80 | 0 |
| 90 | 79/80 | 98.7 | 05/80 | 6.3 | 80/80 | 100 | 7/80 | 8.8 | |
| 15 | 45 | 02/80 | 2.5 | 00/80 | 0 | 01/80 | 1.3 | 0/80 | 0 |
| 90 | 72/80 | 90 | 00/80 | 0 | 75/80 | 93.8 | 3/80 | 3.8 | |
| Crude Extract “ Sonicated ” | Crude Extract “Non-Sonicated” | ||||
| Doses ( kGy ) | Concentrations* | Gamma Radiation | Electron Beams | Gamma Radiation | ElectronBeams |
| 3 | 45 | 70 | 88.8 | 52.5 | 100 |
| 90 | 0 | 40 | 0 | 2.8 | |
| 5 | 45 | 57.5 | 100 | 45 | 90 |
| 90 | 0 | 0 | 0 | 7.5 | |
| 10 | 45 | 97.5 | 100 | 65 | 100 |
| 90 | 1.3 | 1.3 | 0 | 91.3 | |
| 15 | 45 | 97.5 | 100 | 98.8 | 90.6 |
| 90 | 10 | 10 | 6.3 | 96.3 | |
Discussion
Data were Collected through a Combination of the Following Experimental Configurations:
(1) irradiations with gammas and electrons; (2) sonicated and non-sonicated samples; and (3) microcystin concentrations of 45 mg.L-1 and 90 mg.L-1.
Gammas versus Electrons
A cursory visual inspection of Table 1 and Figure 1 shows that the attenuation of microcystin is substantially more efficient following exposure to electron beams, relative to gamma radiation, for both sonicated and non-sonicated cells; that is, Ae >> Aγfor the entire dose interval.
Figure 1.Toxin Attenuation Parameters. Data points: Toxin Attenuation Parameters (Eqn. 1) obtained from irradiations with gammas and electron beams. Curves: Spline drawing of tendency curves as guidelines for the eye.
If electrons as charged particles have quite small penetration lengths in matter, they also display the ability to deposit a large amount of energy in the aqueous medium and molecules, leading to the formation ofsecondary electrons19, 20, 27. As pointed out in section 2.1.2, low-energy secondary electrons have the capacity to inflict damage on DNA and proteins. Secondary electrons are produced by gammas at much smaller amounts.
Our laboratory previously verified this damaging characteristic of electrons 28 and found that plasmids irradiated at intermediate and high doses of electrons are severely shattered into quite small fragments. It is thus clear that toxins exposed to electron beams would be subjected to the same fate of plasmids; they shatter in a rather efficient process to debilitate many of their functions.
Sonicated versus Non-Sonicated Samples
Experiments with non-sonicated and sonicated samples correspond to the irradiation of cells with and without membranes. The implications of irradiation conditions are as follows: (1) innon-sonicated cells (NS), the toxin molecules are confined within the cell interior, where membranes constitute a dense attenuation material for the incoming radiation; (2) in sonicated cells (S), no membrane blocks the incoming radiation, and the toxin molecules are dispersed into the medium.
Table summarizes the results of the Toxin Attenuation Parameters, corresponding to all experimental configurations; note that A(S) > A(NS), for exposure to both gammas and electrons, and with microcystin concentrations of 45 mg.L-1 and 90 mg.L-1. The only exception occurred at 3 kGy for 45 mg.L-1 of microcystin, where A(NS) > A(S) by an amount of only 10%, approximately, but within the range of experimental uncertainties.
The Rationale of these Results is Supported by the Following Considerations
Non-Sonicated Cells
The energy loss as a result of radiation hitting a microcystin molecule occurs by means of (a) interaction with the membrane, and (b) inelastic scattering and rescattering by the clustered material within the cell. Both processes, particularly (a), result in the lowering of the effective dose and therefore imply a smaller attenuation of toxins. Cyanobacteria are known to have rather resistant membranes, proving efficient for radiation energy dissipation29.
(2) Sonicated Cells
With no membrane, the incoming radiation directly hits the cytosol with its full energy, as much as the toxin molecules are dispersed into the aqueous medium. The toxin molecules are not only burdened with direct radiation but also exposed to the total amount of ROS generated by radiolysis. As a consequence, the effective dose imparted to sonicated cells is much higher than in non-sonicated cells, resulting in an equally higher attenuation of toxins, as shown in Table 3.
Microcystin Concentrations: 45 mg.L-1 versus 90 mg.L-1
The results in Table 3show that toxinattenuation for microcystin samples at concentrations of 45 mg.L-1 is much higher than at concentrations of 90 mg.L-1. This is a counterintuitive finding. However, as demonstrated in APPENDIX-A, the average energy absorbed by each target molecule, ε0, is inversely proportional to ρt (mL-1), the target density of toxin molecules; that is, ε0~ 1/ρt. Thus, ε0(45) > ε0(90), implying A(45) > A(90).
Stochasticity of the Attenuation Process
The characteristics of the outcomes (i.e., the number of mobile and immobile individuals) are a consequence of both (1) the Microcystis biophysical conditions and (2) the amount ingested by the Cladocerans.
The Microcystis biophysical conditions refer to the degree of damage inflicted by radiation on its microcystin content, which is a deterministic process, particularly at the doses used in this study. However, the amount of ingested microcystin is a statistically fluctuating quantity, since it is driven by the erratic movement of the Cladocerans in the solution, as described by a simple random walk (similar to a diffusive Brownian motion). Therefore, thevaluesof ingested microcystin at each time is a random variable, thus characterizing the whole process as stochastic.
Deducing Attenuation Functions
The Toxin Attenuation Parameters, Ai(%), were defined in Equation 1, Section 3.4. The stochastic nature of the attenuation process is discussed in Section 4.4. The deduction of a statistically driven expression for Ai presented in APPENDIX-B, Equation B-4, is
Ai(%)=100.(1–exp(–λD))
where the parameter λ is a sole property of the interaction radiation/microcystin. In this case, λ−1 = D0 is the dose necessary to attenuate 63% of the initial microcystin cell population N0.
The data points in Figure 2 and Figure 3 correspond, respectively, to the following experimental conditions: (a) irradiation with gammas, sonicated samples and a concentration of 45 mg/L, and (b) irradiation with electron beams, sonicated samples and concentrations of 45 and 90 mg/L. The curves in Figure 2 and Figure 3 were obtained from a single-parameter best-fitting standard procedure, with λ as the parameter, and imposing Ai = 0 for D = 0. Fitting results are,
(i) λγ = 0.24 ± 0.05 (kGy)-1 (for 45 mg.L-1); therefore (see Figure 2),
Aγ(%)=100.(1–exp(–0.24D)),and
(ii) λe= 0.29 ± 0.04 (kGy)-1 (for 90 mg.L-1); therefore (see Figure 3),
Ae(%)=100.(1–exp(–0.29D)]
Therefore,
D0(e) = 3.45 ± 0.04 (kGy) and D0(γ) = 4.17 ± 0.05 (kGy)
The fact that D0(e) < D0(γ) indicates the higher lethality of electron beams, which is consequence of both (1) their substantial energy transfer to matter and (2) their prolific production of secondary electrons. This is consistent with the present findings of Ae > Aγ.
Figure 2.Irradiation of microcystin with gammas. Data points: Toxin Attenuation Parameter from irradiation of microcystin sonicated samples, concentration of 45 mg.L-1, with gammas. Curve: obtained from the fitting of the expression defined by Eqn. 2 to the data points, resulting in Aγ(%) = 100.1 – exp(–0.24D).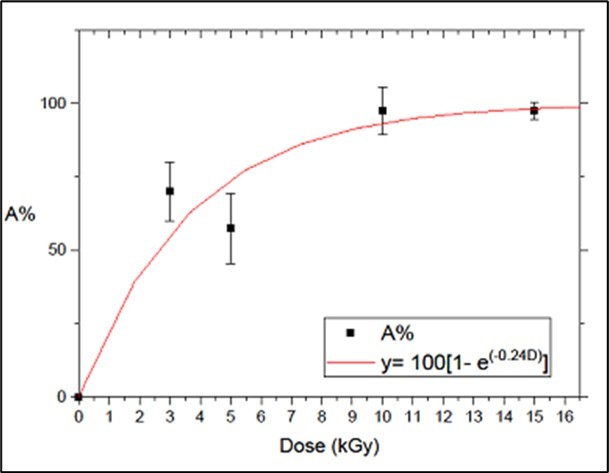
Figure 3.Irradiation of microcystin with electrons. Data points: Toxin Attenuation Parameter from irradiation of microcystin sonicated samples, with electrons at samples concentrations of 45 and 90 mg.L-1. Curve: obtained from the fitting of the expression defined by Eqn. 2 to the data points corresponding to 90 mg.L-1 and resulting in Ae(%) = 100.1 – exp(–0.29D).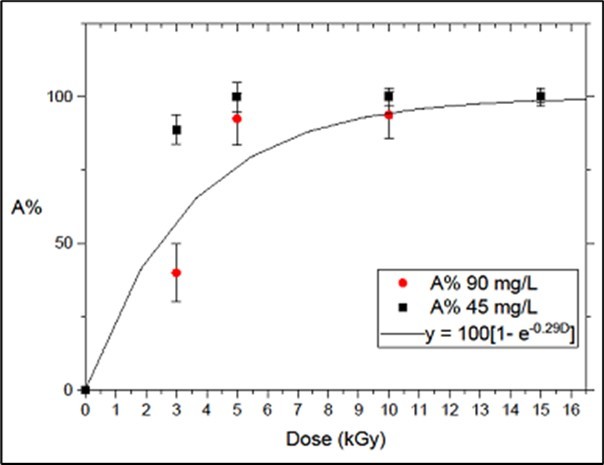
Important Observations
The data handling in Section 4.5 focused on the statistical nature of the results and was carried out with the following data sets: toxin attenuation induced by electron beams in sonicated samples with microcystin concentrations of 90 mg.L-1, and by gammas in sonicated samples with microcystin concentrations of 45 mg.L-1.
The available data were narrowed because (1) compared to electron beams, the energy transfer to the samples by the gamma radiation is extremely low, yielding significant results only for microcystin concentrations of 45 mg.L-1, and sonicated and non-sonicated samples; and (2) attenuation by electrons beams saturates [Ae(%) = 100%] at microcystin concentrations of 45 mg.L-1 for both sonicated and non-sonicated samples.
Application: An Example
In 1998, the city of Miami in the United States, collaborated with the Electron Beam Research Facility (EBRF), also in Miami, to develop and use electron beams for water and wastewater treatment, 30 (see also 31). This facility houses an Electron Linear Accelerator (Linac), powered with an energy of 1.5 MeV (similar to that from a Cobalt source) and an average beam current of 50 mA. The EBRF had already developed excellent know-how for equipment and research protocols and could achieve a water irradiation capability of 460 L per minute.
The feasibility of this EBRF for water/wastewater treatment inspired our laboratory to create a similar device, which is also dedicated to bio-research (as radiobiology and biotechnology). This facility could deliver high-intensity neutron fluxes, another very efficient radiobiological treatment tool (see details in 32).
With completion of the 5 MeV booster, intended for the forthcoming São Paulo Continuous Wave (CW) racetrack microtron 33, a more recent and updated water/wastewater treatment facility is proposed, as outlined in Figure 4. Such a facility could be operated at two irradiation modes: (1) electron beams directly hitting the target (water flux), and (2) electron beams first hitting a radiator, where they produce an intense Bremsstrahlung photon flux on the target.
Figure 4.Water treatment facility. Graphic representation of our proposed water irradiation facility, using electron beams delivered by a 5 MeV-booster from the Continuous Weave (CW) racetrack Microtron at our laboratory 33, similar to a large-scale water/wastewater treatment facility (details in 3031).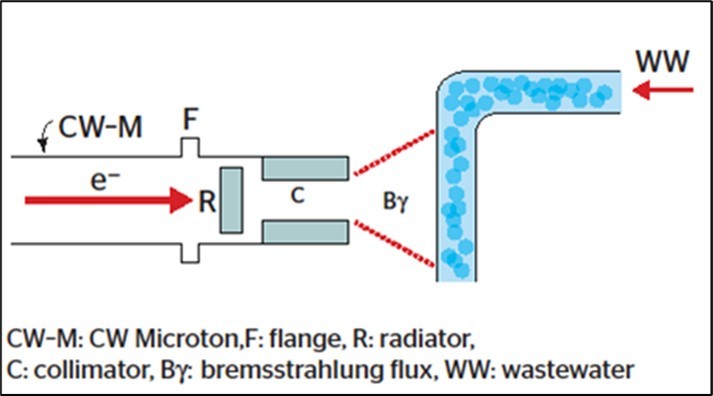
Conclusions
This study yields insights into how to use ionizing radiation to remediate microcystin-contaminated water bodies. This remediation plan could be accomplished from the detailed results here obtained for the radio degradation of microcystin toxicity, as expressed by the attenuation coefficients. The most relevant inferences and conclusions are as follows:
(a) Exposing microcystins to gamma radiation exhibited attenuation at all doses, however in a much smaller and residual proportion than the samples irradiated with electron beams. For microcystin concentrations of 90 mg. L-1, attenuation results for both sonicated and non-sonicated samples were practically null (see Figure 1). Comment: Although the cost of planning and building a gamma irradiation facility is much less than the cost of electron linear accelerator facilities, the cost-benefit ratio is yet high. In addition, the penetrability of gamma rays in microcystin solutions, as limited to microcystin concentrations much smaller than 90 mg.L-1, imposes a veto on the design of large-scale water/wastewater treatment facilities.
(b) Electron beams proved to be powerful physical agents for toxin attenuation, reaching 100% of effectiveness at 5 kGy and above, interpreted as a consequence of the copious amount of low-energy secondary electrons produced by electron beams.
(c) Microcystin attenuation was much higher following irradiations of sonicated samples, for both gammas and electrons, and for microcystin concentrations of 45 mg.L-1 and 90 mg.L-1. Comment: sonicated samples simulate microcystins dissolved in water. Conventional processes are ineffective for removing these extracellular cyanotoxins, as discussed in the Introduction. Incidentally, as shown in Figure 1, at electron doses of 5 kGy and above, attenuation efficiency is approximately 100%.
(d) Counterintuitively, attenuation was higher for microcystins irradiated at a smaller concentration of 45 mg.L-1, a circumstance explained by a biophysical approach worked out in APPENDIX-A. Comment: This is useful information when designing applications in highly contaminated areas.
(e) Saturating doses in the attenuation process were experimentally observed (Figure 1) and conceptually deduced (Eqn. 2). Comment: these are key data for planning and optimizing radiological treatment of water and wastewater.
Table Captions
Table - 1 Mean values of 48 h-EC50, their standard deviations and 95% confidence intervals for the crude extract non-sonicated and sonicated, irradiated with gamma radiation, doses of 3, 5, 10 and 15 kGy for daphinid Ceriodaphniasilvestrii (Crustacea, Cladocera) exposed to extracts of Microcystis aeruginosa (toxic strain NPLJ-4) in acute toxicity tests.
Table - 2 Results of immobility tests of acute toxicity to Ceriodaphnia silvestrii (Crustacea, Cladocera) exposed to extract toxic culture of strain NPLJ 4 of Microcystis aeruginosa, irradiated with electron beams and gamma radiation at doses of 0, 3, 5, 10 and 15 kGy, at concentrations of 45 and 90 mg.L-1 dry weight of freeze-dried material
Table - 3 The Toxin Degradation Parameter, corresponding to data obtained for all the experimental configurations used in this study.
Acknowledgements
The authors acknowledge the Brazilian agencies FAPESP and CNPq for partially supporting this work.
References
- 1.J M O’Neil, T W Davis, M A Burford, C J Gobler. (2012) The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. , Harmful Algae 14, 313-334.
- 2.H W Paerl, N S Hall, E S Calandrino. (2011) Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. , Science of the Total Environment 409, 1739-1745.
- 3.Figueiredo D R de, U M Azeiteiro, S M Esteves, Gonçalves F J M, M J Pereira. (2004) Microcystin-producing blooms: a serious global public health issue. Ecotoxicology and Environmental Safety. 59(2), 151-163.
- 4.R W Zurawell, Chen H, J M Burke, Prepas E E. (2005) Hepatotoxic cyanobacteria: a review of the biological importance of microcystins in freshwater environments. , Journal of Toxicology and Environ mental Health, Part B 8(1).
- 5.M C Bittencourt-Oliveira, M K Cordeiro-Araújo, M A Chia, Arruda-Neto J D T, E T Oliveira et al. (2016) Lettuce irrigated with contaminated water: photosynthetic effects, antioxidative response and bioaccumulation of microcystin congeners. Ecotoxicology and Environmental Safety. 128, 83-90.
- 6.Arruda-Neto J D T, Oliveira M C C, Sarkis J E S, Bordini P, Guevara Manso et al. (2009) Study of environmental burden of lead in children using teeth as bioindicator. , Environment International 35, 614-618.
- 7.Arruda-Neto J D T, L P Geraldo, G R Prado, Garcia F, M C Bittencourt-Oliveira et al. (2010) Study of metals transfer from environment using teeth as biomonitor. , Environment International 36, 243-246.
- 8.E M Jochimsen, W, J S An, D M Cardo, S T Cookson. (1998) Liver failure and death after exposure to microcystins at a hemodialysis center in Brazil. New England. , Journal of Medicine 338, 873-878.
- 9.F M Buratti, Manganelli M, Vichi S, Stefanelli M, Scardala S. (2017) Cyanotoxins: producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. , Archives of Toxicology 91, 1049-1130.
- 10.Campinas M, M J Rosa. (2010) Evaluation of cyanobacterial cells removal and lysis by ultrafiltration. Separation and Purification Technology 70, 345-353.
- 11.Hyenstrand P, Rohrlack T, Beattie A, J S Metcalf, G A Codd et al. (2003) Laboratory studies of dissolved radiolabelled microcystin-LR in lake water. , Water Research 37, 3299-3306.
- 12.Rodríguez E, M E, Meriluoto J, J L Acero. (2007) Oxidation of microcystins by permanganate: reaction kinetics and implications for water treatment. , Water Research 41(1), 102-110.
- 13.Rizzo L. (2011) Bioassays as a tool for evaluating advanced oxidation processes in water and wastewater treatment. , Water Research 45, 4311-4340.
- 14.Mayer-Miebach E, M R Stahl, Eschrig U, Deniaud L, Ehlermann D A E et al. (2005) Inactivation of a non-pathogenic strain of E. coli by ionising radiation. , Food Control 16, 701-705.
- 15.Hossain F, Follett P, K D Vu, Salmieri S, Senoussi C et al. (2014) Radiosensitization of Aspergillus niger and Penicillium chrysogenum using basil essential oil and ionizing radiation for food decontamination. , Food Control 45, 156-162.
- 16.Taghipour F. (2004) Ultraviolet and ionizing radiation for microorganism inactivation. , Water Research 38(18), 3940-3948.
- 17.J E Alpen. (1990) Radiation Biophysics. Prentice-Hall International, Inc. Englewood Cliffs, New Jersey.
- 18.Collin F. (2019) Chemical basis of reactive oxygen species reactivity and involvement in neurodegenerative diseases. , International Journal of Molecular Science 20(10), 2407-2424.
- 19.Sanche L. (2002) Nanoscopic aspects of radio biological damage: fragmentation induced by secondary low-energy electrons. , Mass Spectrometry Reviews 21, 349-369.
- 20.M A Huels, Boudaïffa B, Cloutier P, Hunting D, Sanche L. (2003) Single, double, and multiple double strand breaks induced in DNA by 3-100 eV electrons. , Journal of the American Chemical Society 125, 4467-4477.
- 21.P R Gorham, McLachlan J, U T Hammer, W K Dim. (1964) Isolation and culture of toxic strain of Anabaena flos-aquae (Lyngb.) de Bréb. SIL Proceedings1922–2010 15, 796-804.
- 22.Wang N, C D Ivey, R A Dorman, C G Ingersoll, Steevens J. (2018) Acute Toxicity of Sodium Chloride to a Unionid Mussel in water exposures (Lampsilis Siliquoidea). Environmental Toxicology Chemistry. 37(12), 3041-3049.
- 23.M A Hamilton, R C, R V Thurton. (1977) Trimmed Sperman-Karber method for estimating median lethal concentration in toxicity bioassays. Environmental Science and Technology 11, 714-719.
- 24.Hammer O, Harper D A T, P D Ryan. (2001) Past: Paleontological statistics software package for education and data analysis. , Paleontological Electronic 4, 1-9.
- 25.S I Borrely, A, Oikawa H, C L Duarte, F R Rocha. (2004) Electron beam accelerator for detoxification of effluents. When radiation processing can enhance the acute toxicity. Radiation Physics and Chemistry. 71, 453-456.
- 26.M F Romanelli, Moraes M C F, Villavicencio A L C H, S I Borrely. (2004) Evaluation of toxicity reduction of sodium dodecyl sulfate submitted to electron beam radiation. Radiation Physics and Chemistry. 71, 409-411.
- 27.Denifl S, Sulzer P, Huber D, Zappa F. (2007) Influence of functional groups on the site-selective dissociation of adenine upon low-energy electron attachment. , Angewandte Chemie International Edition 46, 5238-5241.
- 28.Nieto L, Arruda-Neto J D T, M A Cotta, Carrer H, Garcia F. (2012) DNA fragmentation by gamma radiation and electron beams using atomic force microscopy. , Journal of Biological Physics 38, 531-542.
- 29.Hoiczyk E, Hansel A. (2000) Cyanobacterial cell walls: news from an unusual prokaryotic envelope. , Journal of Bacteriology 182(5), 1191-1199.
- 30.C N Kurucz, T D Waite, W J Cooper. (1995) The Miami electron beam research facility: A large scale wastewater treatment application. , Radiation Physics and Chemistry 45, 299-308.
- 31.Siwek M, Edgecock T. (2020) Application of electron beam water radiolysis for sewage sludge treatment a review. , Environmental Science and Pollution Research 27, 42424-42448.