
A Review of Attempts to Improve Cow Fertility Through Reproductive Management: Estrous Synchronisation
Abstract
This review focused on the various methods for controlling estrous cycles in well-managed dairy cows. Because up to 70% of dairy cows may stay non-pregnant after an AI procedure, an effective approach for identifying and reinseminating open cows is essential for dairy herds to achieve optimal reproductive performance. Overall, well-managed dairy farms with effective estrus detection programs inseminate 50% or more of non-pregnant cows after behavioral estrus is detected. Cows not detected in estrus are admitted in a resynchronization of ovulation procedure to receive a timed AI (TAI) service to avoid a long interbreeding interval. In Egypt, a widely used program involves starting the Ovsynch protocol (GnRH-7 d-PGF2-56 h-GnRH-16 to 20 h-TAI) 32 days after an initial AI, regardless of pregnancy status. Previous studies have proven that there was no difference in pregnancy/artificial insemination (P/AI) between Ovsynch+P4 and Presynch-Ovsynch, both protocols were equally effective in improving the fertility of cows with a CL 15 mm. The review also addressed different methods for synchronization of ovulation and different factors affecting the selection of the management program.
Author Contributions
Academic Editor: Yanzhou Yang, Ningxia Medical University
Checked for plagiarism: Yes
Review by: Single-blind
Copyright © 2021 Mohammed Elmetwally, et al.
 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Competing interests
The authors have declared that no competing interests exist.
Citation:
Introduction
The profitability of dairy cows is depending mainly on reproductive performance 1. Reproductive management is considered as one of the most important factors that should be taken into consideration to increase the profitability of dairy cows. Calving interval, milk production efficiency, and herd replacement dynamics, the timing of pregnancy during lactation affect the profitability of dairy herds 2. Although insemination and the risk of conception after the end of the voluntary waiting period (VWP) are the two main determinants of time to pregnancy during lactation, the length of the VWP, which determines when cows are eligible for insemination, can also affect the timing of pregnancy. VWP has traditionally lasted 60 days in dairy farms 3. Dairy farm revenue is affected by maximizing the number of female calves born for substitutions, minimizing replacement due to reproductive disorders, and optimizing the time that adult cows spend in the most efficient part of the lactation curve 1.
Although there are numerous reproductive management strategies available for dairy farms, determining the best management program to implement remains a major challenge for dairy producers owing to the complicated interactions of multiple biological and management factors affecting dairy herd dynamics and economics 4, 5. The dairy industry has recently approved hormonal protocols for synchronizing estrus and ovulation as reproductive management systems to increase the overall reproductive performance of lactating cows 6. Ovsynch protocols are important for increasing insemination risk but not fertility in dairy cattle because the conception rate (CR) of AI services after Ovsynch is typically within the reference range for cows inseminated after observed estrus 4, 7. Recently, new protocols, such as Presynch-Ovsynch and Double-Ovsynch, have recently been developed to improve not only the risk of insemination but also the fertility of timed AI (TAI) 7, 8, 9.
So, the present study aimed to evaluate how the application of different synchronization protocols such as Ovsynch, modified Ovsynch, presynch, and modified presynch protocols would be used to maximize the reproductive performance of Holstein dairy cows in Egypt.
The Estrous Cycle in Dairy Cows
In dairy cows, the estrous cycle is the time between two standing heats. The estrous cycle is a cyclical pattern of ovarian activity that allows female animals to go from a state of reproductive receptivity to one of reproductive non-receptivity, allowing for the formation of pregnancy after mating. An estrous cycle in cattle lasts 18-24 days on average 10, 11. The luteal phase (14-18 days; met-estrus and di-estrus) and the follicular phase (4-6 days; pro-estrus and estrus) are the two distinct periods of the cycle. The luteal phase begins after ovulation when the corpus luteum (CL) is created, and the follicular phase begins when the corpus luteum (CL) dies (luteolysis) and ends with ovulation 11, 12, 13. During the follicular phase, the ovulatory follicle's final maturation and ovulation occur, resulting in the release of an egg (the female gamete) into the oviduct, allowing for fertilization.
GnRH starts a chain reaction of hormones that regulate the estrous cycle 14, 15, 16. Follicle maturation is influenced by FSH, while LH acts on the ovarian tissues at the site of ovulation, changing them into corpus luteum (CL) 15, 17, 18. Furthermore, these hormones control the many stages of estrus, including the follicular phase, estrus, and luteal phase 19, 20, 21, 22. Ovulation is an inflammatory process characterized by an influx of leukocytes into the ovulatory follicle and changes in the translation profile of immune markers in the theca and granulosa tissue layers and has been defined as an inflammatory process 23.
Following ovarian stimulation with FSH and LH, estrogen and progesterone are generated. Because it produces prostaglandin F2, the uterus aids in reproductive control 24, 25, 26.
During the follicular, estrus, and luteal phases of the cycle, the combination of hormone secretion and metabolism maintains the correct hormonal balance 27, 28, 29. The pre-ovulatory follicle and the later formed corpus luteum are the two main ovarian structures that govern the estrous cycle by secreting estradiol and progesterone, respectively 23, 30, 31.
During the ruminant estrous cycle, changes in a pre-ovulatory follicle and corpus luteum, patterns of LH, estrogen, and progesterone release, and changes in ovarian blood flow all occur 32, 33, 34, 35.
Maximizing reproductive performance in high-producing cows is one of the top aims of dairy herd management 36. Oestrus detection failure and artificial insemination timing error may occur in dairy cows with short and weakened estrous signals, resulting in low reproductive success 37. Moreover, the difficulty of detecting oestrus and the resulting low heat detection rate may be major contributors to declining reproductive performance 38, 39.
To attain an appropriate heat detection rate, cows must exhibit estrous symptoms for a significant amount of time, and estrous behaviors must be carefully observed by herdsmen. Due to delayed resumption of ovarian cyclicity and failure of oestrus expression, increased milk production and changes in management techniques can result in a longer duration of post-partum anoestrus 16, 40, 41. The predominant estrous symptom in cattle is standing to be mounted by herd members. Mounting other cows, restlessness, enlargement, relaxation, and congestion of the vulva, clear mucus secretion, bleeding following oestrus, and a decrease in appetite and milk supply are all secondary estrous symptoms in cattle 42, 43, 44. Standing oestrus in dairy cows is reported to last 12-18 hours 45, 46. Estrous detection failure and timing inaccuracy in artificial insemination (AI) might result from shorter oestrus length andweakened estrous signals without standing oestrus, resulting in a low conception rate 37. In the majority of cows with standing oestrus, secondary estrous indicators such as sexual activity and changes to the external genitals were observed. Cows without standing oestrus who were detected in oestrus based on external genital changes as secondary estrous signs had a significantly lower percentage of secondary estrous behavior such as mounting and deviant mounting and chasing when compared to standing estrous cows 37, 47, 48, 49
Of note, the success of the artificial insemination program in most cases depends mainly on the detection of estrous. However, because peak estrus activity generally occurs at night, accurate control of the period of estrus is difficult, and determining the true commencement of standing estrus without 24 hours of observation may be challenging 45, 50
Regular observation of the cows is required for the successful detection of heat. These should be observed at least twice a day, once early in the morning and once later in the evening. At least 20 minutes should be spent on each observation. The more females on heat at the same time, the more heat activity there will be 48, 51, 52. Estrus behavior is suppressed by high stocking rates and concrete or slippery flooring (as opposed to straw yards or grassland).
The optimal time to inseminate a heifer or cow is a few hours before ovulation, which happens about 24-38 hours after standing heat begins. This means cows or heifers should be inseminated in the latter two-thirds of a heat cycle or within a few hours after the cycle has ended 53, 54, 55
The environment (temperature, season, and light), age and body weight, hormonal imbalance, diet, and amount of production are all aspects that influence the expression of estrus behavior in bovine species. The temperature has a significant impact on the expression of estrous in bovines since sexual activity is reduced in extreme cold and hot settings, resulting in poor conception. It also has an impact on the duration of the estrous cycle 48, 56, 57, 58.
The ability to correctly identify cows in estrus and inseminate them at the right time is crucial in reproductive control. In recent decades, technology for recognizing cows in estrus has progressed from tail paint in the 1970s to automated activity monitors and mount detectors 33, 59, 60. The gold standard indicator that a cow is in estrus is standing to be mounted. Cows have been reported to stand for 6 to 10 hours to be mounted, however standing estrus is usually missed due to either weak intensity of expression (cow effect) or insufficient observation (human effect), or both 60. Between the 1990s and the early 2000s, automated estrus detection technologies such as mount detectors, pedometers, and accelerometers were validated and commercially available 61. The effectiveness and accuracy of these devices have been proven, and they provide continuous, automated monitoring and analysis of mounting or moving behavior 50, 62, 63.
Understanding the endocrine and neurological processes that drive estrus in dairy cows makes it easier to devise tactics for increasing estrous behavior identification. A concept map on estrus in cattle has been created (Figure 1), which should be referred to frequently throughout the reading of this work to aid comprehension of the issues covered 64, 65. A preovulatory follicle forms on one of the two ovaries during proestrus. This follicle secretes estrogens in response to pituitary gonadotropins in high amounts, causing plasma concentrations of the major estrogen, estradiol, to reach peak levels 24, 26, 66.
Estrus is caused in the absence of progesterone by the action of estradiol on the hypothalamus (Figure 1). Estradiol has been shown to induce estrus in cattle, horses, sheep, and pigs with ovariectomies. Despite the presence of estrus-inducing serum concentrations of estradiol, vaccination of cattle and sheep against estradiol reduced the development of estrous behavior 24. Estradiol appears to have "all or none" effects in inducing estrus (Figure 1). That is, once the concentration of plasma estradiol is high enough to induce estrus, additional amounts have no effect on the behavior's expression. Additional amounts, however, may be required for reproductive functions, such as the secretion of specific proteins needed to nourish the early embryo 67, 68, 69. In both intact and ovariectomized cattle, this all-or-none effect has been demonstrated 70, 71. Furthermore, when low doses of estradiol are given and 50% of the ovariectomized cattle are induced into estrus, the cows in estrus exhibit a frequency of various behaviors that are similar to that of cows given much higher doses 72, 73.
Figure 1.Concept map of estrus in dairy cattle. MBH=Medical basal hypochamismus, CL= corpus luteum and CRF= cortiocotroin realeasing hormone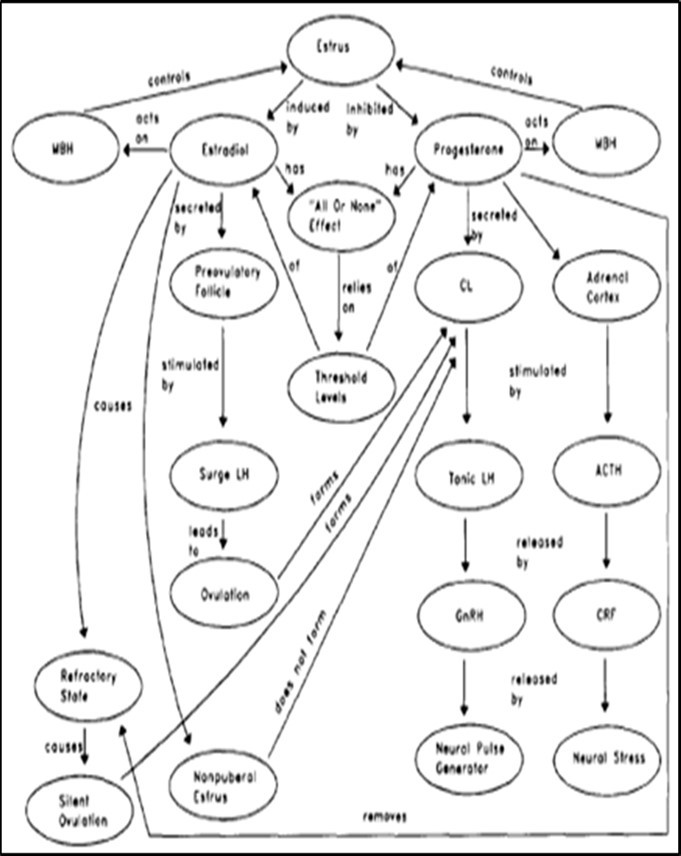
The size of the follicle at the time of GnRH-induced ovulation influenced the formation and maintenance of pregnancy in beef cattle 16, 67, 74. In postpartum beef cows and beef heifers, GnRH-induced ovulation from a tiny dominant follicle (<11.33 mm) was linked with a lower pregnancy rate and late embryonic/fetal survival compared to GnRH-induced ovulation from a large dominant follicle 75, 76, 77.
The lower percentage of cows pregnant after GnRH-induced ovulation from a physiologically immature dominant follicle is likely related to oocyte competence difficulties and insufficient maternal tissue preparation for pregnancy 72, 78. A lower plasma 17-estradiol (E2) concentration before and after ovulation, as well as lower plasma progesterone (P4) concentration after ovulation, may result in insufficient preparation of the maternal tissues for pregnancy 79, 80, 81.
Reproduction Management of Dairy Cows: Estrus Synchronization
Estrus synchronization is a significant reproductive control method in the dairy cattle sector, where the majority of animals are bred through artificial insemination. Estrus synchronization lowers the cost of estrus detection and eliminates mistakes. Manipulation of the length of the luteal phase of the estrus cycle is the core premise of estrus synchronization 82, 83
Although numerous synchronization programs involving one or more hormones have been created and deployed on commercial farms, the fundamental concept of estrus synchronization is to modify the length of the estrus cycle 84, 85. The luteal phase can be shortened or lengthened using prostaglandin F2 or its analogs, and it can also be lengthened with exogenous progestogens. Furthermore, some programs have employed gonadotropin-releasing hormone (GnRH) and estrogens to increase conception rates by modifying follicular wave development and shortening the time between estrus and ovulation 86, 87.
Despite the fact that estrus synchronization is associated with a reduced conception rate during the synchronized estrus, improvements in estrus detection efficiency and accuracy can improve practically all reproductive performance. 87, 88. The fundamental challenge in estrus synchronization research has been to achieve tight synchrony while minimizing the unfavorable effect on the conception rate during the synchronized estrus. Ovulation synchronization is the current trend, which allows for planned breeding without detecting estrus 89.
In fact, despite the advantages of estrus synchronization, commercial dairy producers have yet to implement it. When dairy farmers consider whether or not to use estrus synchronization, they consider the expense of drugs as well as the time it takes to implement the program. Detecting estrus in a large herd of synchronized cattle can be more difficult because so many animals are in estrus at the same time, making it impossible to tell which ones are in real estrus 89, 90, 91. This problem can be solved by developing synchronization routines that allow insemination to occur at a precise time. Herd managers must have strong organizational skills as well as some technical knowledge of the program in order to run an effective estrus synchronization scheme. Herd managers, veterinarians, and AI technicians must all efficiently communicate and collaborate. Because the use of PG on pregnant animals causes abortion, meticulous record-keeping is required 4, 92.
Principles of Estrus Synchronization
Since the introduction of the Ovsynch protocol two decades ago, significant progress has been made in the understanding and application of synchronization programs for the management of reproduction in dairy herds 4, 92. Dairy farmers have quickly adopted scheduled artificial insemination (AI) methods as a result of a better understanding of oestrus cycle regulation and its relationship to inadequate reproductive performance in dairy herds. Fixed-time AI has become an essential component of the management of reproduction in high-producing herds, according to recent assessments 93, 94. Moreover, due to their capacity to improve the insemination rate, timed AI procedures have shown benefits in pasture-based milk production systems. In order to use the Ovsynch protocol successfully, some basic physiological principles must be followed, such as inducing ovulation to synchronize follicle growth in the first two days of the program so that a young antral follicle is recruited; maintaining high progesterone concentrations during the development of the ovulatory follicle while also effectively lysing the ovulatory follicle 95, 96
Furthermore, timed AI protocols have also demonstrated benefits in pasture-based milk production systems because of the ability to increase the insemination rate. In general, successful use of the Ovsynch protocol requires some fundamental physiological principles to be respected, including induction of ovulation to synchronize follicle growth in the first 2 days of the program such that a young antral follicle is recruited; maintenance of high concentrations of progesterone during the development of the ovulatory follicle. Having a good quality pre-ovulatory follicle of moderate diameter that is highly oestrogenic and responsive to gonadotropins to synchronously ovulate 12 to 18 hours after insemination, and having an active corpus luteum to result in very low progesterone concentration at AI, and having a healthy corpus luteum to result in very low progesterone concentration at AI. The present estrous and ovulation synchronization methods are inefficient, and future progress would almost probably demand new hormone composition and delivery technology, lowering the need for intervention and assuring producer acceptance 88.
The dairy industry's consolidation, combined with the steady increase in herd size, has necessitated the implementation of systematic reproduction management programs. The development of synchronized ovulation procedures, which provide artificial insemination at a predetermined period with acceptable fertility, has become an important part of breeding management in a variety of production systems 84. In the last two decades, improvement of these programs has resulted from a greater understanding of ovarian biology in lactating cows, as well as improved control of follicle growth and luteal lifespan, providing unique opportunities to modify follicle development, optimize oocyte quality, and increase embryo survival 97.
Methods of Estrus Synchronization Application in Dairy Cows
For the selection and successful implementation of the estrus synchronization planned, knowledge of the hormonal profile and functional structures present in the ovaries at various stages of the estrous cycle is highly crucial 53, 98. The basic method is to manage the time when the estrus begins by managing the length of the estrous cycle. Different ways to the estrous cycle length control are Prostaglandin shots before natural luteolytic time, for the recovery of the animal's corpus luteum (CL), is by employing gonadotropin release hormones or analogs that cause ovulation of a dominant follicle or ovulation of a larger follicle causing the ovulation of a larger follicle 99, 100.
Prostaglandins Based Protocol Approach
The hormones that exist spontaneously are prostaglandin (PG). PGF will be released from the womb within 16 to 18 days of the heat of the animal during the regular estrous cycle of a non-pregnant animal. This PGF release works to damage the corpus luteum (CL). The CL is an ovarian structure producing the hormone progesterone and avoiding the return of the animal to the estrus. The release of PGF from the uterus causes the animal to return to the estrus every 21 days 101, 102. PGF2α (Lutalyse, Estrumate, Prostamate), which is commercially available, allows the cattle owner the possibility to remove CL concurrently from all the animals at an advantageous moment for warmth detection and raising 53. PGF2α has a key drawback in that it is ineffective in animals that do not have a CL. This comprises animals who have been in heat for 6 to 7 days, prepubertal heifers, and postpartum anestrous cows 103, 104.
Despite these drawbacks, using prostaglandins to synchronize estrus in cattle is the easiest way. These products can synchronize estrus and fertility in cyclic females, such as virgin heifers, but they can't produce estrous cycles in non-cycling cows 105, 106, 107, 108.
One-Shot Prostaglandin F2α
In dairy cows with a functioning corpus luteum, prostaglandin F2 is often utilized to synchronize estrus. When dairy cows are given PGF2, around 50-60% of the treated cows go into estrus within 2-6 days 92, 93, 94. Following artificial insemination after estrous detection, subsequent conception rates have been reported to reach 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42 percent, resulting in a pregnancy rate of around 20% per treated cow 95. As a result, the large time intervals between PGF2 injection and estrus/ovulation, as well as the failure to detect estrus, are important practical drawbacks. To optimize the efficacy of timed AI (TAI), a procedure that closely synchronizes PGF2 therapy and estrus/ovulation is required 50, 84.
Two Shot Prostaglandin
Once the stage of the estrous cycle in the cows is identified, two injections of prostaglandins are administered at 10-to-14-day intervals. Before or between injections, estrus detection is not required. Regardless of where they were in their estrous cycle when the first injection was given, all cycling cows should respond to the second. All females experiencing estrus after the initial PGF2 injection can be bred, which can change the program 109, 110. Then only the females that haven't been bred receive the second injection. This option saves money and time, but it results in two synchronized groups rather than one, and it takes longer to breed.
Recent studies used intravaginal (IVG) prostaglandins instead of intramuscular or subcutaneous. of notes, Wijma et al. (2016) found that IVG delivery of two doses of 25 mg dinoprost (a natural form of PGF2α) 12 hours apart successfully produced complete luteal regression and resulted in circulating progesterone (P4) profile similar to that of milking dairy cows given a single 25-mg IM dosage of PGF2α. Circulating P4 concentrations in cows treated with IVG or IM PGF2α did not differ 48, 60, or 72 hours after therapy, and a similar proportion of cows in both groups exhibited complete CL reduction by 60 and 72 hours following treatment 111. Similarly, were the results for the application of GnRH intravaginal in automated synchronization systems 112.
Wijma et al. (2016) also found that giving two 25-mg dinoprost doses 12 hours apart was the most effective way to get complete CL regression compared to smaller or larger doses (i.e., 25 and 125 mg) or a similar dose (i.e., 50 mg) given as a single therapy. P4 concentration dynamics in cows given two 25-mg PGF2α doses 12 hours apart were similar to those in cows given a single 25-mg IM dosage of dinoprost, suggesting that this dose and treatment frequency for intravaginal infusion is a feasible choice for future investigations examining CL regression and ovulation synchronization 111.
Progesterone Base Protocol Approach
Even after the corpus luteum has regressed, synchronization of estrus with progestogens maintains high amounts of progesterone in the female's system. Estrus synchrony occurs 2 to 5 days after progestin withdrawal. Melengesterol acetate (oral feeding), Syncro-Mate-B (Ear Implant), and CIDR are commercial products that fit within this category (Intra-vaginal device). Only 48% of cows treated on day 3 had their estrus synchronized, but when treatment began on day 9 of the estrous cycle, the synchronization was 100%. The longer cattle were given progestin, the higher the rate of estrous synchronization, but the lower the fertility of the synchronized animals 113, 114, 115, 116. In addition to PGF2α, inserting a progesterone-impregnated intravaginal insert (progesterone-releasing intravaginal device (PRID) or controlled internal drug release (CIDR) insert) during a 5- or 7-day period efficiently synchronizes estrus. When injected at the time of insert removal, PGF2α lyses any functioning corpus luteum. Inseminations usually take place two to five days after estrus is recognized and removed 117, 118.
This therapeutic regimen was used to develop the Syncro-mate B commercial product, as well as the PRID (Progesterone releasing intravaginal device) and CIDR (Controlled intra vaginal drug release device). Although the corpus luteum has shrunk, increased pulsatile secretion of gonadotropin during the period when exogenous progestin is preventing estrus leads the persistent follicle to grow 119. The use of an intravaginal device for 10 to 14 days during any synchronization program increases the risk of local vaginal infection, which, in the worst-case scenario, leads to an ascending infection to the uterus, producing endometritis-pyometra. At the time of CIDR application and withdrawal, strict cleanliness must be kept. For each CIDR application, new latex gloves should be worn, and the applicator should be sanitized between uses. Over the CIDR and applicator, a disinfecting ointment is also applied. When the CIDR devices are removed, the next CIDR is performed with a new glove. When injecting eCG, take care to ensure that the female receives the whole amount 119.
The decreased fertility of cows bred at synchronized estrus after long-term progestin administration is attributed to early ovarian meiosis or aberrant embryo development produced from persisting follicle ova. The use of progestogens in cattle for less than 14 days did not lower the percentage of calves conceived. Furthermore, short-term progestogen exposure induces some anestrus (postpartum or prepubertal) heifers to cycle 120, 121.
GnRH - Based Protocol Approach Synchronization Systems
Ovsynch, a GnRH and PGF2α regimen, was created to synchronize ovulation in dairy calves, with the goal of synchronizing ovulation within an 8-hour interval (from 24 to 32 hours after the second GnRH treatment), allowing TAI without detection of estrus and pregnancy rates of 30 to 40% 4, 122. GnRH was injected on day 0 of the Ovsynch regimen, then by PGF2 after 7 days, and the second infusion of GnRH after 48 hours, with TAI 72 hours following PGF2α. Synchronization with the Ovsynch procedure was successful in detecting estrus, facilitating artificial insemination, and assisting producers in improving reproductive efficiency by causing precise ovulations 92, 122, 4. Although PGF2α is particularly successful at promoting luteolysis from days 6 to 16 of the estrous cycle, the time between therapy and estrus and ovulation is rather variable 123. Early investigations of PGF2α for estrus synchronization were aimed to ensure the presence of a responding CL, either by providing a single PGF2α infusion after palpating a CL per rectum or by giving two PGF2α treatments, 11 or 14 days apart. While fertility in response to a PGF-induced estrus has been shown to be comparable to that of a spontaneous estrus, pregnancy outcomes in PGF-based protocols using timed-AI (TAI; without estrus detection) have been considered unacceptably low 124. Timed artificial insemination (TAI) following ovulation synchronization is a technology that is critical for reproductive success in many dairy and beef cow enterprises in North America and around the world 4, 92, 125. Synchronization of ovulation procedures entails using a sequence of two or more hormonal therapies to control follicular wave dynamics, corpus luteum (CL) regression, and ovulation 4. In the dairy cows industry, breeders can choose from a variety of simple hormonal treatment protocols that vary in form, number, and sequence, all with the goal of assuring quick insemination after the voluntary waiting period or a failure in AI treatment 126. Finally, ovulation timing is synchronized within a short time frame, allowing for insemination of groups of cows by a fixed time, regardless of estrus expression and detection 127.
Fertility to AI in dairy heifers is frequently regarded as the gold standard to obtain in lactating dairy cows, and that such fertile responses in heifers should be achieved in TAI programs for both heifers and lactating cows. Lactating dairy cows have a number of problems, including the biological responses and demands of lactation, metabolism, and production disorders include metabolic, mammary, and uterine diseases, all of which reduce fertility. Due to basic reproductive differences, however, developing a fertile TAI program in heifers has proven difficult. Application of a simple OvSynch-program48 in the early stages of TAI development 81, 128, 129
Presynch techniques that include gonadotropin-releasing hormone (GnRH) and prostaglandin (PG) F2 induce ovulation in anovulatory cattle and increase the proportion of cows that synchronize during an Ovsynch procedure maximize pregnancies per A.I. In a seven-day Ovsynch protocol, adding a second PGF2α injection 24 hours after the first boost luteal regression, which leads to more pregnancies per A.I. especially in multiparous cows 130, 9, 131.
The intervals between PGF2α therapy and estrus and ovulation relied on the stage of development of the dominant follicle at the time of treatment, according to preliminary research involving estrus monitoring and following comprehensive ultrasound analyses of ovarian follicular wave patterns 122, 124. PGF2α given 5 or 8 days after ovulation caused the dominant follicle of the first follicular wave to ovulate in 2 or 3 days, but PGF given 12 days after ovulation caused the dominant follicle of the subsequent follicular wave to ovulate in 5 days 132.
There was a rise in the number of synchronized animals and a reduction in the variability in the time to estrus in beef cows and heifers when a GnRH analog was administered 6 days before injection of PGF2. This reduction in time variability could be explained by the commencement of a new follicular wave in response to GnRH, resulting in the presence of a new dominant follicle at the time of PGF2α injection 4, 92.
GnRH-induced follicle turnover or the initiation of a new follicular wave would be most useful if ovulation was prompted in response to the first administration of GnRH, resetting follicular development, and producing a new dominant follicle containing at least 25.8% of the dairy cattle in the Ovsynch group had insufficient luteal regression 133, 134, 135.
Pregnant Mare Serum Gonadotropins (PMSG) and Estrous Synchronization
Late embryonic and early fetal mortality, which average 13 percent and 11 percent, respectively, impair breastfeeding dairy cows' reproductive effectiveness 136, 137. In breastfeeding dairy cows, low plasma progesterone levels are a primary cause of poor embryo development and higher pregnancy losses. In fact, cows with a higher number of corpora lutea than embryos (extra corpus luteum) were less likely to lose a fetus. As a result, increasing progesterone levels in plasma by inducing auxiliary corpora lutea or administering progesterone decreased late embryonic and early fetal mortality 138, 139.
In Bos indicus cows, intervention with intravaginal P4 devices coupled with eCG at device removal enhanced ovulation rates, plasma P4 concentrations, and pregnancy rates in suckled beef cattle with a higher incidence of anestrous or a poor body condition score 140, 141, 142.
The efficiency of TAI procedures is limited in postpartum anestrous cows because their pulsatile release of LH is insufficient to support the final phases of ovarian follicular growth and ovulation 143. Exogenous progestins caused ovulation by increasing LH pulse frequency during and after treatment 144. Despite progestin-induced postpartum cyclicity, the efficacy of such treatment may be hampered in populations with a high number of anestrous cattle, poor body condition scores, or both 144 In order to boost LH support, gonadotropins could be added in synchronization methods.
Because of its cost-effectiveness, pregnant mare serum gonadotropin (PMSG) is commonly used for estrus synchronization programs in small and large ruminants 141, 145. PMSG enhances follicular development and ovulation in cattle by acting like FSH but also like LH. Its parenteral injection stimulates follicular growth and ovulation in cattle 146, 147. PMSG is given to cows either the ovaries produce a pool of little follicles or cause ovulation of big follicles. Following that, PGF2 was given to help with corpus luteum regression. On day 14 postpartum, PMSG treatment improves follicular development and plasma estradiol levels in cattle without influencing subsequent reproductive function 147.
Treatment with PMSG is thought to improve ovarian follicular development and fertility in dairy cows at the end of the estrus synchronization program 122, 148, 149, 150. In the dairy cows' estrous cycle, on Day 4 of the first follicular wave of the estrous cycle, when LH receptors are first discovered in granulosa cells of the dominant follicle 151, maturing ovarian follicles have the highest amount of FSH receptors. After gaining LH receptors in granulosa cells, dominant follicles react to both FSH and LH or eCG 151. In nulliparous beef heifers, injection of eCG at the same time as PGF2 increased the size of the pre-ovulatory ovarian follicle 143 and raised circulating estradiol levels in suckled cattle. On day 14 postpartum, PMSG treatment improves follicular development and plasma estradiol levels in cattle without influencing subsequent reproductive function 122, 148. PMSG is used to either increase the number of small follicles or triggers ovulation in large follicles on the ovaries, followed by PGF2 to reduce corpus luteum growth 152.
Detection of Oestrus is Suboptimal in Dairy Cows
The rate of submission to insemination and P/AI define the rate of pregnancy in dairy herds once the voluntary waiting period has ended. The reduced rate of oestrus detection and, as a result, reduced submission to insemination found in nursing dairy cows is one of the constraints to optimal reproduction in many dairy farms 84, 122. Cows are usually eligible for insemination when they have completed the voluntary waiting period and are not pregnant. Because the oestrus cycle of dairy cows is estimated to last 21 days, appropriate cows should show oestrus and be eligible for insemination every three weeks 85. As a result, the proportion of eligible cows inseminated for each 21-day interval out of the total eligible cattle in the same period is commonly used to compute oestrus diagnosis. Because oestrus identification is the only way to inseminate cows, pregnancy rates decrease when compared to management programs that allow for systematic AI control 4, 92, 153. The delaying in first postpartum insemination and the longer period between re insemination in non-pregnant cows are to account for the reduction in the conception rate 154.
Cows should be Chosen Based on the following Criteria Sufficient time has passed between calving and the start of synchronization treatments; a minimum of 40 days postpartum is recommended at the start of therapy. On a scale of 1 to 9, a body condition score of at least 5 is considered average or above average. Calving issues are minimal 155.
Replacement to avoid issues during synchronization, heifers are grown to a pre-breeding target weight of at least 65 percent of their predicted mature weight, and reproductive tract scores of 2 or higher on a scale of 1 to 5 are assigned to heifers two weeks before synchronization treatment begins 156, 157. To improve the likelihood of responding to a synchronization program, heifers must reach puberty before being synchronized. In addition, compared to her pubertal estrus, a heifer's third estrus results in a 21% increase in fertility 158, 159.
Factors Affecting Estrus Synchronization
Several factors have been proven to alter the length of the estrous cycle, including age, species, and body weight, amount of nourishment, time of the year, hormonal changes, lactation, suckling, and degree of milk yield 160. The effects of herd conflict and stress on the estrous cycle are well established. The duration of oestrus in the farmyard is likely to vary depending on breed, management, and a variety of environmental factors: The average duration of the hot spell is 12-16 hours, with a larger range of 3-28 hours seen across the board 65, 160.
Ovulation has been observed to occur 10-12 hours after the heat cycle has ended 4, 26, 161. Because sperm cells have a short life in the female reproductive system, the moment of ovulation is critical if maximal breeding success is to be achieved 162. The research findings on the timing of ovulation in cattle have been inconsistent, and the number of animals employed in the studies has been limited. According to Foot 163, ovulation happens spontaneously in cows between 24 and 48 hours following the commencement of estrus, based on his own research and that of others. Ovulation occurs 27 hrs from the commencement or just after the conclusion of estrus 37, 164
Body Condition of the Animal
On a scale of one to five, a score of one suggests that the cow is malnourished, while a score of five means that the cow is overweight. A BCS of 3 indicates that the animal has adequate energy stores to support a pregnancy around the time of breeding; a BCS of less than 3 suggests that the animal does not have enough energy reserves to maintain a pregnancy. Because the cow has mobilized body fat stores to fulfill the energy requirements for milk yield in early lactation, negative feeling balance (NEB) is indicated by a drop in BCS 165, 166.
The state of a cow's body has a direct impact on one's ability to reproduce. In dairy cows, there is a lot of evidence that body condition affects reproductive performance. Low body condition can have a detrimental impact on female fertility for a variety of reasons, including a prolonged interval between parturition and the commencement of ovarian activity 167. The single most critical element determining when beef heifers and cows will begin cycling after calving is their body condition at the time of calving 168.
Undernutrition can affect pituitary activity, which includes gonadotropin production and release, as well as the responses of target organs to gonadotropin-releasing hormone or gonadal hormones. The capacity of an animal to maintain a high-frequency mode of pulsatile LH release has been related to its metabolic status. One important way that energy limitation impairs reproductive activity appears to be suppression of an increase in luteinizing hormone pulse the frequency that has been necessary for the growth of ovarian follicles inhibits pulsatile release by the hypothalamus 167, 169, 170.
The Parity Effects
The time between calving and first ovulation in primiparous cows has been shown to be longer than in multiparous cows. Because of the requirements for growth other than lactation, this link is related to more nutritional shortages being imposed on young cows. According to recent research, primiparous cows' first ovulation postpartum was postponed under optimal management compared to multiparous cows. 171.
Multiparous cows have superior reproductive performance than primiparous cows 93. Others discovered that primiparous cows performed equally well or better than multiparous cows. Reduced incidence of metabolic problems in early lactation could be one factor for primiparous cows' better fertility94.
Breed of the Cows’ Effects
The breed plays an important role in the response of cows to the synchronization program. There are considerable differences in the effects of estrus synchronization between zebu and taurine breeds due to their different estrus cycles. There has been documented diversity in the length, duration, and severity of the estrus cycle within and across breeds 172, 173, 174. Estrus lasts 4.8 and 7.4 hours in zebu and their crossbreeds, respectively. Crossbreeding has also been demonstrated to improve estrus appearance and allow calves to reach puberty 6 to 12 months longer than Bos taurus cows 175.
Recently, slight changes in ovarian follicular dynamics that could affect the use of assisted reproduction like synchronization or superovulation have been addressed. 176. Because of the differences in the estrus cycle or the duration of the estrus cycle, synchronization techniques for one do not always work for the other171
Lactation/Suckling and Level of Milk Production
Because prolactin function peaks during intense lactation, and sensory cues from sucking decrease the prolactin inhibiting factor (PIF), which comprises dopamine and GnRH-related peptide, lactation has been demonstrated to diminish ovarian activity. Because dopamine and GnRH have essential interactions with gonadotropins, the GnRH component is important. Because the LH releasing factor is blocked, no LH is released, and hence no final follicle maturation, oestrus, or ovulation occurs 177, 178. When compared to low-producing cows, high-producing cows have worse oestrus behavior and a shorter oestrus duration. It's been claimed that heavy producer cows don't have oestrus cycles for 3 to 4 months after giving birth 87, 178.
Stress
Even in the presence of estrus-inducing doses of estradiol, stress has been shown to delay, shorten, or entirely block the manifestation of estrus in cows 179. The proper operation of the cow's endocrine system is known to be influenced by several forms of stress, whether caused by poor feeding, management, or environmental conditions 31, 173. Many ordinary husbandry procedures can cause a rise in plasma glucocorticoids, and stress in the cow can result in elevated concentrations of progesterone of adrenal origin. Increased cortisol release has been observed in cattle in response to a number of acute stresses 180, 181.
The Influence of Estrous Synchronization Protocol on the Conception Rates in Dairy Cows
Breeding at Fixed-Time Artificial Insemination (FTAI)
Larson and Ball (1992) found that pregnancy rates from fixed TAI after synchronization with PGF2 were varied, especially in lactating dairy cows compared to heifers, owing to changes in the time of ovulation in response to AI, resulting in varying estrous cycle duration in cows 182. Estrus detection accuracy is a critical management issue 183. Approximately half of all estrous episodes go undetected. As a result, the number of insemination possibilities is extremely limited, resulting in increased days open and financial loss. Because of inaccuracies in estrous identification, many cows are inseminated at periods when pregnancy is unlikely 184.
Pursley et al. (1995 and 1997) developed Ovsynch to build a TAI program that put less emphasis on detecting estrus because all cows were inseminated at a specific time relative to hormone injection 92, 4. According to some sources, a fixed-time insemination scheme could revolutionize dairy cow reproductive management by eliminating the requirement for estrus detection 89, 94, 134. Others found that cows getting AI at observed estrus after GnRH-PGF2 had higher conception rates than those receiving Ovsynch with the FTAI program 185.
TAI methods minimized the annoyances of ovulation synchronization and AI on detected estrus, and they suggested that they could provide an efficient and effective means of capturing selecting genetic features with economic implications in a fixed time insemination program. They also noted that changes in pasture and diet, breed composition, physical condition, postpartum interval, climate, and geographic location will impact TAI protocol performance 186. All of these obstacles have prompted scientists to develop a number of estrous synchronization regimens with TAI that are tailored to the needs of each region.
Conception Rate
The initial service pregnancy rate was a useful metric for assessing fertility, with 60-70 % being considered ideal in well-managed herds 187. Several previous studies have looked at the fertility of postpartum dairy cows using the Ovsynch procedure, with fertility rates ranging from 27 to 39 percent each AI 4, 188, 189. Pursley et al. (1995) found that when PGF2 was given 48 and 24 hours before the second GnRH, the rates of conception were 55 and 46 percent, respectively. They also discovered that pregnancy rates varied from 37, 40, 44, 40, and 32 percent in cows inseminated at fixed times of 0, 8, 16, 24, and 32 hours following the second injection of GnRH in the Ovsynch procedure 4. Burke et al. (1996) examined the efficiency of TAI and AI in nursing dairy cows with identified estrus after the Ovsynch protocol and found that conception rates were 30.5 and 29.0 percent, respectively 185.
Progesterone and Estrogen Changes and Estrous Synchronization
The progesterone hormone is in charge of stimulating cyclicity, follicular growth, and pregnancy maintenance. The plasma protein, cholesterol, and mineral profiles of animals indicate their nutritional status and are associated with their fertility 190, 191. Cholesterol, as a major contributor of steroid hormones, is important in steroidogenesis, while calcium tones up the genitalia, and protein and inorganic phosphorus are implicated in metabolic pathways at the cellular level. Several metabolic and environmental factors disrupt these hormonal and nutritional statuses, impairing normal physiology in the animal body 191.
PGF2 therapy resulted in a decrease in progesterone levels on day 9 (48 hours later) in all controlled breeding programs (Ovsynch and CIDR synch) due to PGF2's luteolytic action on the corpus luteum 191, 192.
Significant increase in plasma P4 profile observed on day 7 of treatments with CIDR and Ovsynch protocols (4.97 ±1.68 and 3.75 ±0.47 ng/ml) over initial (0 days) values, with dramatic fall to almost basal values on induced estrus within 48-60 h after PG injection has previously been identified in anestrus cows using CIDR and Ovsynch protocols 190, 193.
Some researchers reported lower first service conception rates (CR) in targeted breeding programs compared to spontaneous oestrus breeding 194. Reducing the estrous cycle repeatedly may interrupt corpus luteum function and reduce fertility in high-yielding dairy cows. Rosenberg et al. (1990) hypothesized that the amount of progesterone produced during the cycle preceding artificial insemination could influence conception 195. Ovulatory failure has been blamed for dairy cows' inability to develop luteal tissue after synchronization 196, 197. The significance of follicular wave dynamics in the onset of oestrus and ovulation timing after synchronization was proven 198
Additionally, serum P4 levels are higher 14 days later than they were at the time of the first treatment. At any given time, a certain percentage of cycling cows have physiologically low progesterone levels. During lactation, the percentage of cows with consistently low P4 levels in serum (1 ng/ml) decreased. Inactive ovaries or cystic ovarian disease are associated with low progesterone levels at PG shots of the ovsynch protocol 194.
Regarding the level of estrogen (E2) during the ovsynch programs, it was reported that GnRH-induced ovulation of small follicles (11.3 mm) in beef cows was associated with lower P/AI and serum E2 concentrations at AI, as well as higher pregnancy loss 199. Moreover, in cows, proestrus estradiol concentration is positively related to the size of the preovulatory follicle, subsequent CL size, serum P4 concentration, and, as a result, pregnancy rate 200. of notes, on days 0 and 7, estrogen levels in various animals treated with Ovsynch protocols were nearly identical 200.
References
- 1.Giordano J O, Fricke P M, Wiltbank M C. (2011) An economic decision-making support system for selection of reproductive management programs on dairy farms. , J Dairy Sci 94, 6216-6232.
- 2.Giordano J O, Kalantari A S, Fricke P M. (2012) A daily herd Markov-chain model to study the reproductive and economic impact of reproductive programs combining timed artificial insemination and estrus detection. , J Dairy Sci 95, 5442-5460.
- 3.Miller R H, Norman H D, Kuhn M T. (2007) Voluntary waiting period and adoption of synchronized breeding in dairy herd improvement herds. , J Dairy Sci 90, 1594-1606.
- 4.Pursley J R, Mee M O, Wiltbank M C. (1995) Synchronization of ovulation in dairy cows using PGF2α and GnRH. Theriogenology. 44, 915-923.
- 5.Souza A H, Ayres H, Ferreira R M. (2008) A new presynchronization system (Double-Ovsynch) increases fertility at first postpartum timed AI in lactating dairy cows. Theriogenology. 70, 208-215.
- 6.Olynk N J, Wolf C A. (2008) Economic analysis of reproductive management strategies on US commercial dairy farms. , J Dairy Sci 91, 4082-4091.
- 7.Fricke P M, Giordano J O, Valenza A. (2014) Reproductive performance of lactating dairy cows managed for first service using timed artificial insemination with or without detection of estrus using an activity monitoring system. , J Dairy Sci 97, 2771-2781.
- 8.Voelz B E, Rocha L, Scortegagna F. (2018) Response of lactating dairy cows with or without purulent vaginal discharge to gonadotropin-releasing hormone and prostaglandin F2α. J Anim Sci. 96, 56-65.
- 9.Stangaferro M L, Wijma R W, Giordano J O. (2019) Profitability of dairy cows submitted to the first service with the Presynch-Ovsynch or Double-Ovsynch protocol and different duration of the voluntary waiting period. , J Dairy Sci 102, 4546-4562.
- 10.LOE Silva, Valenza A, RLOR Alves. (2021) Progesterone release profile and follicular development in Holstein cows receiving intravaginal progesterone devices. , Theriogenology 172, 207-215.
- 11.Perez Marquez HJ, Ambrose D J, Schaefer A L. (2021) Evaluation of infrared thermography combined with behavioral biometrics for estrus detection in naturally cycling dairy cows. Animal. 15, 100205.
- 12.JCL Motta, Madureira G, Silva L O. (2020) Interactions of circulating estradiol and progesterone on changes in endometrial area and pituitary responsiveness to GnRH†. Biol Reprod. 103, 643-653.
- 13.Veronese A, Marques O, Peñagaricano F. (2019) Genomic merit for reproductive traits. II: Physiological responses of Holstein heifers. , J Dairy Sci 102, 6639-6648.
- 14.Savio J D, Thatcher W W, Badinga L. (1993) Regulation of dominant follicle turnover during the oestrous cycle in cows. , J Reprod Fertil 97, 197-203.
- 15.Webb R, Nicholas B, Gong J G. (2003) Mechanisms regulating follicular development and selection of the dominant follicle. , Reprod; 61, 71-90.
- 16.Elmetwally M A. (2018) Uterine involution and ovarian activity in postpartum holstein dairy cows. A review. JVHC. 1, 29-40.
- 17.Webb R, Campbell B K, Garverick H A. (1999) Molecular mechanisms regulating follicular recruitment and selection. J Reprod Fertil Suppl. 54, 33-48.
- 18.Riccetti L, Sperduti S, Lazzaretti C. (2018) The cAMP/PKA pathway: steroidogenesis of the antral follicular stage. Minerva Ginecol. 70, 516-524.
- 19.Luo X, Chang H-M, Yi Y. (2021) Bone morphogenetic protein 2 upregulates SERPINE2 expression through noncanonical SMAD2/3 and p38 MAPK signaling pathways in human granulosa-lutein cells. , FASEB J 35-21845.
- 20.Meier S, Kuhn-Sherlock B, Amer P A. (2021) Positive genetic merit for fertility traits is associated with superior reproductive performance in pasture-based dairy cows with seasonal calving. , J Dairy Sci 104, 10382-10398.
- 21.Basavaraja R, Drum J N, Sapuleni J. (2021) Down regulated luteolytic pathways in the transcriptome of early pregnancy bovine corpus luteum are mimicked by interferon-tau in vitro. , BMC Genomic 22, 452.
- 22.Mahé C, Zlotkowska A M, Reynaud K. (2021) Sperm migration, selection, survival, and fertilizing ability in the mammalian oviduct†. , Biol Reprod 105, 317-331.
- 23.Abdulrahman Alrabiah N, ACO Evans, Fahey A G. (2021) Immunological aspects of ovarian follicle ovulation and corpus luteum formation in cattle. , Reproduction 162, 209-225.
- 24.Ginther O J. (2021) Switching of follicle destiny so that the second largest follicle becomes dominant in monovulatory species. , Theriogenology 171, 147-154.
- 25.Garcia-Ispierto I, Llobera-Balcells M, López-Gatius F. (2021) Inducing ovulation with human chorionic gonadotrophin improves the pregnancy rate in lactating dairy cows receiving an in vitro-produced embryo. Reprod Domest Anim. 56, 1145-1147.
- 26.F Pereira de Moraes, Amaral D’Avila C, F Caetano de Oliveira. (2021) Prostaglandin F2α regulation and function during ovulation and luteinization in cows. , Theriogenology 171, 30-37.
- 27.Piccinno M, Sciorsci R L, Masciopinto V. (2017) Effects of d-cloprostenol on different layers and regions of the bovine uterus during the follicular and luteal phases. , Theriogenology 96, 92-96.
- 28.Sakaguchi K, Yanagawa Y, Yoshioka K. (2019) Relationships between the antral follicle count, steroidogenesis, and secretion of follicle-stimulating hormone and anti-Müllerian hormone during follicular growth in cattle. Reprod Biol Endocrinol. 17-88.
- 29.Nishimura R, Okuda K. (2020) Multiple roles of hypoxia in bovine corpus luteum. , J Reprod Dev 66, 307-310.
- 30.Oosthuizen N, Cooke R F, Schubach K M. (2020) Effects of estrous expression and intensity of behavioral estrous symptoms on variables associated with fertility in beef cows treated for fixed-time artificial insemination. Anim Reprod Sci. 214-106308.
- 31.Sakai S, Yagi M, Fujime N. (2021) Heat stress influences the attenuation of prostaglandin synthesis by interferon tau in bovine endometrial cells. , Theriogenology 165, 52-58.
- 32.EOS Batista, Vieira L M, Freitas B G. (2020) Anti-Mullerian hormone and its relationship to ovulation response and fertility in timed AI Bos indicus heifers. Reprod Domest Anim. 55, 753-758.
- 33.Schweinzer V, Gusterer E, Kanz P. (2020) Comparison of behavioral patterns of dairy cows with natural estrus and induced ovulation detected by an ear-tag based accelerometer. , Theriogenology 157, 33-41.
- 34.Ramachandran R, Vinothkumar A, Sankarganesh D. (2020) Detection of estrous biomarkers in the body exudates of Kangayam cattle (Bos indicus) from interplay of hormones and behavioral expressions. Domest Anim Endocrinol. 72, 106392.
- 35.Schweinzer V, Gusterer E, Kanz P. (2019) Evaluation of an ear-attached accelerometer for detecting estrus events in indoor housed dairy cows. , Theriogenology. 130: 19-25
- 36.López-Gatius F. (2003) Is fertility declining in dairy cattle? A retrospective study in northeastern Spain. , Theriogenology 60, 89-99.
- 37.Yoshida C, Nakao T. (2005) Some characteristics of primary and secondary oestrous signs in high-producing dairy cows. , Reprod Domest Anim 40, 150-155.
- 38.Neglia G, D de Nicola, Esposito L. (2020) Reproductive management in buffalo by artificial insemination. , Theriogenology; 150, 166-172.
- 39.Kunii H, Kubo T, Asaoka N. (2021) Loop-mediated isothermal amplification (LAMP) and machine learning application for early pregnancy detection using bovine vaginal mucosal membrane. , Biochem Biophys Res Commun; 569, 179-186.
- 40.Steeneveld W, Hogeveen H. (2015) Characterization of Dutch dairy farms using sensor systems for cow management. , J Dairy Sci 98, 709-717.
- 41.Richardson B N, Hill S L, Stevenson J S. (2016) Expression of estrus before fixed-time AI affects conception rates and factors that impact expression of estrus and the repeatability of expression of estrus in sequential breeding seasons. Anim Reprod Sci. 166, 133-140.
- 42.García-Guerra A, Kirkpatrick B W, Wiltbank M C. (2017) Follicular waves and hormonal profiles during the estrous cycle of carriers and non-carriers of the Trio allele, a major bovine gene for high ovulation and fecundity. , Theriogenology 100, 100-113.
- 43.Díaz R, Galina C S, Rubio I. (2017) Resumption of ovarian function, the metabolic profile and body condition in Brahman cows (Bos indicus) is not affected by the combination of calf separation and progestogen treatment. , Anim Reprod Sci 185, 181-187.
- 44.Zhang S, Mao W, Li Q. (2018) Concentration effect of prostaglandin E2 on the growth factor expression and cell proliferation in bovine endometrial explants and their kinetic characteristics. Reprod Domest Anim. 53, 143-151.
- 45.Cunha T O, Martinez W, Walleser E. (2021) Effects of GnRH and hCG administration during early luteal phase on estrous cycle length, expression of estrus and fertility in lactating dairy cows. , Theriogenology; 173, 23-31.
- 46.Held-Montaldo R, Cartes D, Sepúlveda-Varas P. (2021) Behavioral changes in dairy cows with metritis in seasonal calving pasture-based dairy system. , J Dairy Sci
- 47.Fricke P M, Carvalho P D, Giordano J O.Expression and detection of estrus in dairy cows: the role of new technologies. , Animal8Suppl1: 134-143.
- 48.AML Madureira, Silper B F, Burnett T A. (2015) Factors affecting expression of estrus measured by activity monitors and conception risk of lactating dairy cows. , J Dairy Sci 98, 7003-7014.
- 49.Reith S, Hoy S. (2018) Review: Behavioral signs of estrus and the potential of fully automated systems for detection of estrus in dairy cattle. Animal. 12, 398-407.
- 50.Tippenhauer C M, Plenio J L, AML Madureira. (2021) Factors associated with estrous expression and subsequent fertility in lactating dairy cows using automated activity monitoring. , J Dairy Sci 104, 6267-6282.
- 51.Marques O, Veronese A, Merenda V R. (2020) Effect of estrous detection strategy on pregnancy outcomes of lactating Holstein cows receiving artificial insemination and embryo transfer. , J Dairy Sci 103, 6635-6646.
- 52.Burnett T A, Polsky L, Kaur M. (2018) Effect of estrous expression on timing and failure of ovulation of Holstein dairy cows using automated activity monitors. , J Dairy Sci 101, 11310-11320.
- 53.Wood S L, Lucy M C, Smith M F. (2001) Improved synchrony of estrus and ovulation with the addition of GnRH to a melengestrol acetate-prostaglandin F2alpha synchronization treatment in beef heifers. , J Anim Sci 79, 2210-2216.
- 54.Meng Chao L, Sato S, Yoshida K. (2010) Comparison of oestrous intensity between natural oestrus and oestrus induced with ovsynch based treatments in Japanese black cows. Reprod Domest Anim. 45, 168-170.
- 55.Denis-Robichaud J, LeBlanc S J, Jones-Bitton A. (2018) Pilot study to evaluate the association between the length of the luteal phase and estrous activity detected by automated activity monitoring in dairy cows. Front Vet Sci. 5-210.
- 56.Schindler H, Eger S, Davidson M. (1991) Factors affecting response of groups of dairy cows managed for different calving-conception intervals. Theriogenology. 36, 495-503.
- 57.Yániz J L, Santolaria P, Giribet A. (2006) Factors affecting walking activity at estrus during postpartum period and subsequent fertility in dairy cows. Theriogenology. 66, 1943-1950.
- 58.AML Madureira, Polsky L B, Burnett T A. (2019) Intensity of estrus following an estradiol-progesterone-based ovulation synchronization protocol influences fertility outcomes. , J Dairy Sci 102, 3598-3608.
- 59.Stevenson J S, Britt J H. (2017) A 100-Year Review: Practical female reproductive management. , J Dairy Sci 100, 10292-10313.
- 60.Moore S G, Aublet V, Butler S T. (2021) Monitoring estrous activity in pasture-based dairy cows. , Theriogenology 160, 90-94.
- 61.Stevenson J S, Hill S L, Nebel R L. (2014) Ovulation timing and conception risk after automated activity monitoring in lactating dairy cows. , J Dairy Sci 97, 4296-4308.
- 62.Tippenhauer C M, Plenio J L, AML Madureira. (2021) Timing of artificial insemination using fresh or frozen semen after automated activity monitoring of estrus in lactating dairy cows. , J Dairy Sci 104, 3585-3595.
- 63.Schilkowsky E M, Granados G E, Sitko E M. (2021) Evaluation and characterization of estrus alerts and behavioral parameters generated by an ear-attached accelerometer-based system for automated detection of estrus. , J Dairy Sci 104, 6222-6237.
- 64.Cardinali D P, Vacas M I, Lowenstein P R. (1979) [Neurohumoral control of the pineal gland. A model for the study of neuroendocrine integrative processes]. Acta Physiol Lat Am. 29, 291-304.
- 65.Allrich R D. (1994) Endocrine and neural control of estrus in dairy cows. , J Dairy Sci 77, 2738-2744.
- 66.Etchevers L, Belotti E M, Díaz P U.2021; MC2R/MRAP2 activation could affect bovine ovarian steroidogenesis potential after ACTH treatment. , Theriogenology 174, 102-113.
- 67.Manuel Palomino J, Grand F-X, Vigneault C. (2020) Effects of follicular ablation and GnRH on synchronization of ovulation and conception rates in embryo recipient heifers. Anim Reprod Sci. 221-106596.
- 68.Bai Y, Zhang F, Zhang H. (2020) Follicular Fluid Metabolite Changes in Dairy Cows with Inactive Ovary Identified Using Untargeted Metabolomics. Biomed Res Int. 9837543.
- 69.Agarwal S, Gupta H P, Prasad S. (2021) Effect of various hCG treatment protocols on luteal characteristics, plasma progesterone concentration, and pregnancy in normal cyclic Indian crossbred dairy cows. Trop Anim Health Prod. 53-220.
- 70.Ray D E. (1965) Oestrous response of ovariectomized beef heifers to oestradiol benzoate and human chorionic gonadotrophin. , J Reprod Fertil 10, 329-335.
- 71.Walton J S, Veenhuizen L P, King G J. (1987) Relationships between time of day, estrous behavior, and the preovulatory luteinizing hormone surge in Holstein cows after treatment with cloprostenol. , J Dairy Sci 70, 1652-1663.
- 72.Jinks E M, Smith M F, Atkins J A. (2013) Preovulatory estradiol and the establishment and maintenance of pregnancy in suckled beef cows. , J Anim Sci 91, 1176-1185.
- 73.Ciernia L A, Perry G A, Smith M F. (2021) Effect of estradiol preceding and progesterone subsequent to ovulation on proportion of postpartum beef cows pregnant. Anim Reprod Sci. 227-106723.
- 74.Stevenson J S. (2019) Spatial relationships of ovarian follicles and luteal structures in dairy cows subjected to ovulation synchronization: Progesterone and risks for luteolysis, ovulation, and pregnancy. , J Dairy Sci 102-5686.
- 75.de Carvalho NAT, de Carvalho JGS, de Souza DC. (2021) Lack of effect of melatonin on ovarian function and response to estrous synchronization and fixed-time AI during the nonbreeding season in lactating dairy buffalo (Bubalus bubalis). Anim Reprod Sci. 231-106796.
- 76.Garcia-Ispierto I, Pando M, Llobera-Balcells M. (2021) Inducing Ovulation with hCG Improves Fertility Outcomes of Co-Dominant Follicle Drainage to Avoid Twin Pregnancy in Dairy Cows. Animals (Basel). 11.
- 77.Mahdavi-Roshan H, Niasari-Naslaji A, Vojgani M. (2020) Size and number of corpora lutea and serum progesterone concentrations when administering two doses of eCG in an estrous synchronization treatment regimen for dairy cattle. Anim Reprod Sci. 222-106620.
- 78.Atkins J A, Smith M F, Wells K J. (2010) Factors affecting preovulatory follicle diameter and ovulation rate after gonadotropin-releasing hormone in postpartum beef cows. Part II: Anestrous cows. , J Anim Sci 88, 2311-2320.
- 79.Macmillan K L, BVE Segwagwe, Pino C S. (2003) Associations between the manipulation of patterns of follicular development and fertility in cattle. Anim Reprod Sci. 78, 327-344.
- 80.Hori K, Matsuyama S, Nakamura S. (2019) Age-related changes in the bovine corpus luteum function and progesterone secretion. Reprod Domest Anim. 54, 23-30.
- 81.AML Madureira, Burnett T A, Borchardt S. (2021) Plasma concentrations of progesterone in the preceding estrous cycle are associated with the intensity of estrus and fertility of Holstein cows. PLoS ONE. 16-0248453.
- 82.Knight T W, Lambert M G, Devantier B P. (2001) Calf survival from embryo transfer-induced twinning in dairy-beef cows and the effects of synchronised calving. , Anim Reprod Sci 68, 1-12.
- 83.Osawa T, Honjou S, Nitta H. (2009) Effect of synchronisation of ovulation on ovarian profile and days open in holstein cows diagnosed as nonpregnant 26 days after timed artificial insemination. J Reprod Dev. 55, 163-169.
- 84.Masello M, Ceglowski B, Thomas M J. (2020) A reproductive management program aimed at increasing reinsemination of nonpregnant dairy cows at detected estrus resulted in similar reproductive performance to a program that favored timed artificial insemination. , J Dairy Sci 103, 3719-3729.
- 85.Pérez M M, Wijma R, Scarbolo M. (2020) Lactating dairy cows managed for second and greater artificial insemination services with the Short-Resynch or Day 25 Resynch program had similar reproductive performance. , J Dairy Sci 103, 10769-10783.
- 86.Lee M-S, Rahman M S, Kwon W-S. (2013) Efficacy of four synchronization protocols on the estrus behavior and conception in native Korean cattle (Hanwoo). Theriogenology. 80, 855-861.
- 87.Carvalho P D, Santos V G, Giordano J O. (2018) Development of fertility programs to achieve high 21-day pregnancy rates in high-producing dairy cows. , Theriogenology 114, 165-172.
- 88.Giordano J O, Thomas M J, Catucuamba G. (2016) Reproductive management strategies to improve the fertility of cows with a suboptimal response to resynchronization of ovulation. , J Dairy Sci 99, 2967-2978.
- 89.Lopes G, Johnson C R, LGD Mendonça. (2013) Evaluation of reproductive and economic outcomes of dairy heifers inseminated at induced estrus or at fixed time after a 5-day or 7-day progesterone insert-based ovulation synchronization protocol. , J Dairy Sci 96, 1612-1622.
- 90.Busch W, Klinsky J D, Slucka R. (1977) [Planned control of reproduction processes in industrialized cattle production]. Arch Exp Veterinarmed. 31, 799-808.
- 91.Cavalieri J, Macmillan K L. (2002) Synchronisation of oestrus and reproductive performance of dairy cows following administration of oestradiol benzoate or gonadotrophin releasing hormone during a synchronised pro-oestrus. Aust Vet J. 80, 486-493.
- 92.Pursley J R, Wiltbank M C, Stevenson J S. (1997) Pregnancy rates per artificial insemination for cows and heifers inseminated at a synchronized ovulation or synchronized estrus. , J Dairy Sci 80, 295-300.
- 93.Thomas J M, Poock S E, Ellersieck M R. (2014) Delayed insemination of non-estrous heifers and cows when using conventional semen in timed artificial insemination. , J Anim Sci 92, 4189-4197.
- 94.Abel J M, Bishop B E, Thomas J M. (2017) Comparing strategies to synchronize estrus before fixed-time artificial insemination in primiparous 2-year-old beef cows. Theriogenology. 87, 306-315.
- 95.Jeong J K, Kang H G, Hur T Y. (2013) Synchronization using PGF(2α) and estradiol with or without GnRH for timed artificial insemination in dairy cows. , J Reprod Dev 59, 97-101.
- 96.JEP Santos, Bartolome J A, RLA Cerri. (2004) Effect of a deslorelin implant in a timed artificial insemination protocol on follicle development, luteal function and reproductive performance of lactating dairy cows. Theriogenology. 61, 421-435.
- 97.Bilby T R, RGS Bruno, Lager K J. (2013) Supplemental progesterone and timing of resynchronization on pregnancy outcomes in lactating dairy cows. , J Dairy Sci 96, 7032-7042.
- 99.Overton T R, JAA McArt, Nydam D V. (2017) A 100-Year Review: Metabolic health indicators and management of dairy cattle. , J Dairy Sci 100, 10398-10417.
- 100.Moore S G, Hasler J F. (2017) A 100-Year Review: Reproductive technologies in dairy science. , J Dairy Sci 100, 10314-10331.
- 101.Borchardt S, Haimerl P, Heuwieser W. (2016) Effect of insemination after estrous detection on pregnancy per artificial insemination and pregnancy loss in a Presynch-Ovsynch protocol: A meta-analysis. , J Dairy Sci 99, 2248-2256.
- 102.Bonacker R C, Stoecklein K S, JWC Locke. (2020) Treatment with prostaglandin F2α and an intravaginal progesterone insert promotes follicular maturity in advance of gonadotropin-releasing hormone among postpartum beef cows. , Theriogenology 157, 350-359.
- 103.de Souza LB, Dupras R, Mills L. (2013) Effect of synchronization of follicle-wave emergence with estradiol and progesterone and superstimulation with follicle-stimulating hormone on milk estrogen concentrations in dairy cattle. , Can J Vet Res 77, 75-78.
- 104.AML Madureira, Burnett T A, Pohler K G. (2020) Short communication: Greater intensity of estrous expression is associated with improved embryo viability from superovulated Holstein heifers. , J Dairy Sci 103, 5641-5646.
- 105.Stevenson J S, Hoffman D P, Nichols D A. (1997) Fertility in estrus-cycling and noncycling virgin heifers and suckled beef cows after induced ovulation. , J Anim Sci 75, 1343-1350.
- 106.Wright P J, Malmo J. (1992) Pharmacologic manipulation of fertility. Vet Clin North Am Food Anim Pract. 8, 57-89.
- 107.Kohram H, Poorhamdollah M. (2012) Relationships between the ovarian status and superovulatory responses in dairy cattle. , Anim Reprod Sci 131, 123-128.
- 108.Sato T, Nakada K, Uchiyama Y. (2005) The effect of pretreatment with different doses of GnRH to synchronize follicular wave on superstimulation of follicular growth in dairy cattle. , J Reprod Dev 51, 573-578.
- 109.Machado V S, Neves R C, Lima F S. (2017) The effect of Presynch-Ovsynch protocol with or without estrus detection on reproductive performance by parity, and the long-term effect of these different management strategies on milk production, reproduction, health and survivability of dairy cows. Theriogenology. 93, 84-92.
- 110.Noronha I M, Cooke R F, CFG Martins. (2020) Administering an additional prostaglandin F2α injection to Bos indicus beef cows during a treatment regimen for fixed-time artificial insemination. Anim Reprod Sci. 219-106535.
- 111.Wijma R, Stangaferro M L, Giordano J O. (2016) Circulating progesterone dynamics after intravaginal instillation of prostaglandin-F2α to lactating dairy cows. Theriogenology. 85, 1660-1668.
- 112.Wijma R, Stangaferro M L, Masello M. (2017) Intravaginal instillation of gonadotropin-releasing hormone analogues with an absorption enhancer induced a surge of luteinizing hormone in lactating dairy cows. , J Dairy Sci 100, 7626-7637.
- 113.Sahu S K, Cockrem J F, Parkinson T J. (2015) Effects of progesterone inclusion in a gonadotropin-prostaglandin-gonadotropin programme on follicular dynamics and ovulation synchronisation of pasture-based dairy cows with anoestrous. , Res Vet Sci 102, 200-205.
- 114.SAA Edwards, Boe-Hansen G B, Satake N. (2015) A field investigation of a modified intravaginal progesterone releasing device and oestradiol benzoate based ovulation synchronisation protocol designed for fixed-time artificial insemination of Brahman heifers. , Anim Reprod Sci 160, 105-111.
- 115.Sahu S K, Cockrem J F, Parkinson T J. (2017) Effects of GnRH, a progesterone-releasing device, and energy balance on an oestrus synchronisation program in anoestrous dairy cows. Aust Vet J. 95, 281-288.
- 116.JJ de la Mata, Núñez-Olivera R, Cuadro F. (2018) Effects of extending the length of pro-oestrus in an oestradiol- and progesterone-based oestrus synchronisation program on ovarian function, uterine environment and pregnancy establishment in beef heifers. Reprod Fertil Dev. 30, 1541-1552.
- 117.AA-A Swelum, Saadeldin I M, Moumen A. (2018) Efficient follicular wave synchronization using a progesterone-releasing intravaginal device (PRIDΔ) in Camelus dromedarius. , Theriogenology 118, 203-211.
- 118.Macan R C, Camargo C E, Zielinski B L. (2021) Timed artificial insemination in crossbred mares: Reproductive efficiency and costs. Reprod Domest Anim. 56, 459-466.
- 119.Fernandez-Novo A, Santos-Lopez S, Pesantez-Pacheco J L. (2021) . Effects on Synchronization and Reproductive Efficiency of Delaying the Removal of the Intravaginal Progesterone Device by 24 h in the 5d Co-Synch Protocol in Heifers. Animals , (Basel) 11.
- 120.T van Werven, Waldeck F, Souza A H. (2013) Comparison of two intravaginal progesterone releasing devices (PRID-Delta vs CIDR) in dairy cows: blood progesterone profile and field fertility. Anim Reprod Sci. 138, 143-149.
- 121.SAA Edwards, Atkinson P C, Satake N. (2014) Ovarian dynamics in response to two modified intravaginal progesterone releasing device and oestradiol benzoate based ovulation synchronisation protocols designed for use in Brahman heifers. Anim Reprod Sci. 148, 18-25.
- 122.Hussien A, Sharawy H, Lenis Y. (2021) The impact of different estrus synchronization programs on postpartum holstein dairy cow reproductive performance. Mans Vet Med J. 22, 124-130.
- 123.Roche J F. (1974) Synchronization of oestrus and fertility following artificial insemination in heifers given prostaglandin F2 alpha. , J Reprod Fertil 37, 135-138.
- 124.Martínez M F, Kastelic J P, Mapletoft R J. (2004) The use of estradiol and/or GnRH in a two-dose PGF protocol for breeding management of beef heifers. Theriogenology. 62, 363-372.
- 125.Yousuf M R, JPN Martins, Ahmad N. (2016) Presynchronization of lactating dairy cows with PGF2α and GnRH simultaneously, 7 days before Ovsynch have similar outcomes compared to G6G. Theriogenology. 86, 1607-1614.
- 126.Wijma R, Pérez M M, Masello M. (2018) A resynchronization of ovulation program based on ovarian structures present at nonpregnancy diagnosis reduced time to pregnancy in lactating dairy cows. , J Dairy Sci 101, 1697-1707.
- 127.Ruelle E, Shalloo L, Butler S T. (2021) Economic impact of different strategies to use sex-sorted sperm for reproductive management in seasonal-calving, pasture-based dairy herds. , J Dairy Sci
- 128.Randi F, Kelly A K, Parr M H. (2021) Effect of ovulation synchronization program and season on pregnancy to timed artificial insemination in suckled beef cows. , Theriogenology 172, 223-229.
- 129.Kasimanickam R, Kasimanickam V, Kappes A. (2021) Timed artificial insemination strategies with or without short-term natural service and pregnancy success in beef heifers. , Theriogenology 166, 97-103.
- 130.Karakaya-Bilen E, Yilmazbas-Mecitoglu G, Keskin A. (2019) Fertility of lactating dairy cows inseminated with sex-sorted or conventional semen after Ovsynch, Presynch-Ovsynch and Double-Ovsynch protocols. Reprod Domest Anim. 54, 309-316.
- 131.Stevenson J S, Sauls J A, LGD Mendonça. (2018) Dose frequency of prostaglandin F2α administration to dairy cows exposed to presynchronization and either 5- or 7-day Ovsynch program durations: Ovulatory and luteolytic risks. , J Dairy Sci 101, 9575-9590.
- 132.Kaneko K, Mungthong K, Noguchi M. (2020) Day of prostaglandin F2α administration after natural ovulation affects the interval to ovulation, the type of ovulated follicle, and the failure to induce ovulation in cows. , J Vet Med Sci 82, 590-597.
- 133.Zuluaga J F, Saldarriaga J P, Cooper D A. (2010) Presynchronization with gonadotropin-releasing hormone increases the proportion of Bos indicus-influenced females ovulating at initiation of synchronization but fails to improve synchronized new follicular wave emergence or fixed-time artificial insemination conception rates using intravaginal progesterone, gonadotropin-releasing hormone, and prostaglandin F2alpha1. J Anim Sci. 88, 1663-1671.
- 134.Perry G A, Perry B L, Krantz J H. (2012) Influence of inducing luteal regression before a modified fixed-time artificial insemination protocol in postpartum beef cows on pregnancy success. , J Anim Sci 90, 489-494.
- 135.Satheshkumar S, Subramanian A, Devanathan T G. (2012) Follicular and endocrinological turnover associated with GnRH induced follicular wave synchronization in Indian crossbred cows. Theriogenology. 77, 1144-1150.
- 136.JEP Santos, Thatcher W W, Chebel R C. (2004) The effect of embryonic death rates in cattle on the efficacy of estrus synchronization programs. Anim Reprod Sci. 82(83), 513-535.
- 137.López-Gatius F, Santolaria P, Yániz J. (2002) Factors affecting pregnancy loss from gestation Day 38 to 90 in lactating dairy cows from a single herd. Theriogenology. 57, 1251-1261.
- 138.Thatcher W W, Moreira F, Pancarci S M. (2002) Strategies to optimize reproductive efficiency by regulation of ovarian function. Domest Anim Endocrinol. 23, 243-254.
- 139.López-Gatius F, Santolaria P, Yániz J L. (2004) Progesterone supplementation during the early fetal period reduces pregnancy loss in high-yielding dairy cattle. Theriogenology. 62, 1529-1535.
- 140.Baruselli P S, Reis E L, Marques M O. (2004) The use of hormonal treatments to improve reproductive performance of anestrous beef cattle in tropical climates. , Anim Reprod Sci 82(83), 479-486.
- 141.NAT Carvalho, Soares J G, Porto Filho RM. (2013) Equine chorionic gonadotropin improves the efficacy of a timed artificial insemination protocol in buffalo during the nonbreeding season. Theriogenology. 79, 423-428.
- 142.Sá Filho MF, JRS Torres-Júnior, Penteado L. (2010) Equine chorionic gonadotropin improves the efficacy of a progestin-based fixed-time artificial insemination protocol in Nelore (Bos indicus) heifers. Anim Reprod Sci. 118, 182-187.
- 143.Sá Filho MF, Ayres H, Ferreira R M. (2010) Equine chorionic gonadotropin and gonadotropin-releasing hormone enhance fertility in a norgestomet-based, timed artificial insemination protocol in suckled Nelore (Bos indicus) cows. Theriogenology. 73, 651-658.
- 144.Meneghetti M, Sá Filho OG, RFG Peres. (2009) Fixed-time artificial insemination with estradiol and progesterone for Bos indicus cows I: basis for development of protocols. Theriogenology. 72, 179-189.
- 147.Kamomae H, Kaneda Y, Domeki I. (1989) Blood LH level and induced ovulation after hCG and PMSG treatment in ovarian quiescent cattle. Nippon Juigaku Zasshi. 51, 467-473.
- 148.Souza A H, Viechnieski S, Lima F A. (2009) Effects of equine chorionic gonadotropin and type of ovulatory stimulus in a timed-AI protocol on reproductive responses in dairy cows. Theriogenology. 72, 10-21.
- 150.Pulley S L, Wallace L D, Mellieon H I. (2013) Ovarian characteristics, serum concentrations of progesterone and estradiol, and fertility in lactating dairy cows in response to equine chorionic gonadotropin. Theriogenology. 79, 127-134.
- 151.Ireland J J, Roche J F. (1982) Development of antral follicles in cattle after prostaglandin-induced luteolysis: changes in serum hormones, steroids in follicular fluid, and gonadotropin receptors. Endocrinology. 111, 2077-2086.
- 152.Sheldon I M, Dobson H. (2000) Effect of administration of eCG to postpartum cows on folliculogenesis in the ovary ipsilateral to the previously gravid uterine horn and uterine involution. , J Reprod Fertil 119, 157-163.
- 153.Middleton E L, Pursley J R. (2019) Short communication: Blood samples before and after embryonic attachment accurately determine non-pregnant lactating dairy cows at 24 d post-artificial insemination using a commercially available assay for pregnancy-specific protein B. , J Dairy Sci 102, 7570-7575.
- 154.Tenhagen B A, Drillich M, Surholt R. (2004) Comparison of timed AI after synchronized ovulation to AI at estrus: reproductive and economic considerations. , J Dairy Sci 87, 85-94.
- 155.Meier S, Kay J K, Kuhn-Sherlock B. (2020) Effects of far-off and close-up transition cow feeding on uterine health, postpartum anestrous interval, and reproductive outcomes in pasture-based dairy cows. , J Anim Sci Biotechnol 11-17.
- 156.Schmidt C, Gajewski Z, Wehrend A. (2013) [Strategic hormonal reproductive programs in cows. Part 1: overview, Ovsynch and its modifications]. Tierarztl Prax Ausg G Grosstiere Nutztiere. 41, 45-54.
- 157.El-Zarkouny S Z, Shaaban M M, Stevenson J S. (2011) Blood metabolites and hormone-based programmed breeding treatments in anovular lactating dairy cows. , J Dairy Sci 94, 6001-6010.
- 158.Roth Z, Kressel Y Z, Lavon Y. (2021) Administration of GnRH at Onset of Estrus, Determined by Automatic Activity Monitoring, to Improve Dairy Cow Fertility during the Summer and Autumn. Animals (Basel). 11.
- 159.Ataide Junior GA, Kloster A, de Moraes ÉG. (2021) Early resynchronization of follicular wave emergence among Nelore cattle using injectable and intravaginal progesterone for three timed artificial inseminations. Anim Reprod Sci. 229-106759.
- 160.Shiferaw Y, Tenhagen B A, Bekana M. (2005) Reproductive disorders of crossbred dairy cows in the central highlands of Ethiopia and their effect on reproductive performance. Trop Anim Health Prod. 37, 427-441.
- 161.Bó G A, Baruselli P S, Chesta P M. (2006) The timing of ovulation and insemination schedules in superstimulated cattle. Theriogenology. 65, 89-101.
- 162.Walker W L, Nebel R L, McGilliard M L. (1996) Time of ovulation relative to mounting activity in dairy cattle. , J Dairy Sci 79, 1555-1561.
- 163.Foote R H. (1996) Review: dairy cattle reproductive physiology research and management—past progress and future prospects. , J Dairy Sci 79, 980-990.
- 164.Nowicki A, Barański W, Baryczka A. (2017) OvSynch Protocol and its Modifications in the Reproduction Management of Dairy Cattle Herds - an Update. , J Vet Res 61, 329-336.
- 165.Scipioni R L, Foote R H, Lamb S V. (1982) Electronic probe measurements of cervico-vaginal mucus for detection of ovulation in dairy cows: sanitation, clinical observations and microflora. Cornell Vet. 72, 269-278.
- 166.Talukder S, Kerrisk K L, Ingenhoff L. (2014) Infrared technology for estrus detection and as a predictor of time of ovulation in dairy cows in a pasture-based system. Theriogenology. 81, 925-935.
- 167.Cardoso Consentini CE, Wiltbank M C, Sartori R. (2021) Factors That Optimize Reproductive Efficiency in Dairy Herds with an Emphasis on Timed Artificial Insemination Programs. Animals (Basel). 11.
- 168.Roche J R, Friggens N C, Kay J K. (2009) Invited review: Body condition score and its association with dairy cow productivity, health, and welfare. , J Dairy Sci 92, 5769-5801.
- 169.Alkan H, Karaşahin T, Dursun Ş. (2020) Evaluation of the factors that affect the pregnancy rates during embryo transfer in beef heifers. Reprod Domest Anim. 55, 421-428.
- 170.Tríbulo P, Balzano-Nogueira L, Conesa A. (2019) Changes in the uterine metabolome of the cow during the first 7 days after estrus. Mol Reprod Dev. 86, 75-87.
- 171.Bayemi P H, Leinyuy I, Nsongka M V. (2015) Effect of cow parity and synchronization method with PGF2α on conception rates of Bos indicus cows in Cameroon. Trop Anim Health Prod. 47, 159-162.
- 172.Forro A, Tsousis G, Beindorff N. (2015) Factors affecting the success of resynchronization protocols with or without progesterone supplementation in dairy cows. , J Vet Sci 16, 121-126.
- 173.Macmillan K L. (2010) Recent advances in the synchronization of estrus and ovulation in dairy cows. , J Reprod Dev 56, 42-7.
- 174.Bisinotto R S, Ribeiro E S, JEP Santos.Synchronisation of ovulation for management of reproduction in dairy cows. Animal 2014; 8 Suppl 1, 151-159.
- 175.Burnett T A, AML Madureira, Silper B F. (2017) Integrating an automated activity monitor into an artificial insemination program and the associated risk factors affecting reproductive performance of dairy cows. , J Dairy Sci 100, 5005-5018.
- 176.Nogueira É, Silva AS da, Amaral T B. (2012) Follicular dynamics and production of oocytes in young Nellore heifers with energetic supplementation. R Bras Zootec. 41, 2012-2017.
- 177.Niozas G, Tsousis G, Steinhöfel I. (2019) Extended lactation in high-yielding dairy cows. I. Effects on reproductive measurements. , J Dairy Sci 102, 799-810.
- 178.Lucy M C. (2001) Reproductive loss in high-producing dairy cattle: where will it end?. , J Dairy Sci 84, 1277-1293.
- 179.Dobson H, Routly J E, Smith R F. (2020) Understanding the trade-off between the environment and fertility in cows and ewes. Anim Reprod. 17-20200017.
- 180.Nanda A S, Dobson H, Ward W R. (1990) Relationship between an increase in plasma cortisol during transport-induced stress and failure of oestradiol to induce a luteinising hormone surge in dairy cows. Res Vet Sci. 49, 25-28.
- 181.Dobson H, Smith R F. (2000) What is stress, and how does it affect reproduction? Anim Reprod Sci. 60(61), 743-752.
- 182.Larson L L, Ball P J. (1992) Regulation of estrous cycles in dairy cattle: A review. Theriogenology. 38, 255-267.
- 183.Ball P J, Cowpe J E, Harker D B. (1983) Evaluation of tail paste as an oestrus detection aid using serial progesterone analysis. , Vet Rec 112, 147-149.
- 184.Guner B, Erturk M, Yilmazbas-Mecitoglu G. (2020) Effect of delaying the time of insemination with sex-sorted semen on pregnancy rate in Holstein heifers. Reprod Domest Anim. 55, 1411-1417.
- 185.Burke J M, sal Sota RL, CA de la Risco. (1996) Evaluation of timed insemination using a gonadotropin-releasing hormone agonist in lactating dairy cows. , J Dairy Sci 79, 1385-1393.
- 186.Lamb G C, Stevenson J S, Kesler D J. (2001) Inclusion of an intravaginal progesterone insert plus GnRH and prostaglandin F2alpha for ovulation control in postpartum suckled beef cows. , J Anim Sci 79, 2253-2259.
- 187.Yildiz A. (2010) Effect of administering ovsynch protocol plus postbreeding infusionon first service pregnancy outcome in cows. , J of Animal and Veterinary Advances 9, 1345-1350.
- 188.Bisinotto R S, Castro L O, Pansani M B. (2015) Progesterone supplementation to lactating dairy cows without a corpus luteum at initiation of the Ovsynch protocol. , J Dairy Sci 98, 2515-2528.
- 189.Lima F S, Bisinotto R S, Ribeiro E S. (2012) Effect of one or three timed artificial inseminations before natural service on reproductive performance of lactating dairy cows not observed for detection of estrus. Theriogenology. 77, 1918-1927.
- 190.Nak Y, Tuna B, Nak D. (2011) The effects of ovsynch, ovsynch with progestin and progestin plus double TAI on pregnancy rates in unobserved oestrus dairy cows and heifers. Kafkas Üniversitesi Veteriner Fakültesi Dergisi.
- 191.Dhami A J, Nakrani B B, Hadiya K K. (2015) Comparative efficacy of different estrus synchronization protocols on estrus induction response, fertility and plasma progesterone and biochemical profile in crossbred anestrus cows. Vet World. 8, 1310-1316.
- 192.Bhoraniya H L, Dhami A J, Naikoo M. (2012) Effect of estrus synchronization protocols on plasma progesterone profile and fertility in postpartum anestrous Kankrej cows. Trop Anim Health Prod. 44, 1191-1197.
- 193.Kumar L, Phogat J B, Pandey A K. (2016) Estrus induction and fertility response following different treatment protocols in Murrah buffaloes under field conditions. Vet World. 9, 1466-1470.
- 194.Tenhagen B A, Birkelbach E, Heuwieser W. (2000) Serum progesterone levels in post-partum dairy cows after repeated application of the prostaglandin F2 alpha analogue D (+) cloprostenol sodium. J Vet Med A Physiol Pathol Clin Med. 47, 213-220.
- 195.Rosenberg M, Kaim M, Herz Z. (1990) Comparison of methods for the synchronization of estrous cycles in dairy cows. 1. Effects on plasma progesterone and manifestation of estrus. , J Dairy Sci 73, 2807-2816.
- 196.Mohammadi A, Seifi H A, Farzaneh N. (2019) Effect of prostaglandin F2α and GnRH administration at the time of artificial insemination on reproductive performance of dairy cows. Vet Res Forum. 10, 153-158.
- 197.Uslu B A, Kocyigit A, Sendag S. (2020) The effect of GnRH on the pregnancy ratio in low-yielding local race cows: comparison of different injection times. Trop Anim Health Prod. 52, 497-502.
- 198.FCF Dias, Costa E, Adams G P. (2013) Effect of duration of the growing phase of ovulatory follicles on oocyte competence in superstimulated cattle. Reprod Fertil Dev. 25, 523-530.
Cited by (5)
This article has been cited by 5 scholarly works according to:
Citing Articles:
N. Mariol, Atef B. Mahdy, Hussein A. Amer, Ali A. Aghwider, A. Hazzaa - Open Veterinary Journal (2024) Semantic Scholar
K. B. Putro, A. Amrozi, Adi Winarto, A. Boediono, Wasmen Manalu - Jurnal Ilmu Ternak dan Veteriner (2024) Semantic Scholar
Reproduction in Domestic Animals (2023) OpenAlex
Ovozoa Journal of Animal Reproduction (2022) OpenAlex
Ovozoa : Journal of Animal Reproduction (2022) Crossref
Dian Mulfristia, H. Saputra, Cut Nila Thasmi, Hafizuddin Hafizuddin, Ginta Riady et al. - Ovozoa : Journal of Animal Reproduction (2022) Semantic Scholar